 Труженик сельского хозяйства
Труженик сельского хозяйства| Труженик сельского хозяйства |
|
РАЗДЕЛ II Основные типы повреждений плодовых и ягодных культур низкими отрицательными температурамиРастения смородины характеризуются высокой потенциальной морозоустойчивостью, повреждаются или вымерзают они только в случае выращивания в условиях избыточной влажности почвы, засухи, неравномерного или ограниченного увлажнения либо же поражения листьев антракнозом, листьев и побегов мучнистой росой, а также при иных функциональных нарушениях. ПОВРЕЖДЕНИЕ МОРОЗАМИ КОРНЕВОЙ СИСТЕМЫ Корневая система плодовых деревьев характеризуется низкой морозоустойчивостью. Так, если надземная часть устойчивых сортов яблони зимой, при благоприятных условиях подготовки к зиме, может перенести, не повреждаясь, температуру минус 41—42 °С и ниже, то корневая система большинства плодовых культур повреждается при температуре минус 10—12°, иногда минус 16 °С. Степень морозоустойчивости корней плодовых деревьев непостоянна в течение осенне-зимнего и зимне-весеннего периодов. В эти периоды изменяется также последовательность в устойчивости отдельных тканей. Если зимой корни плодовых деревьев повреждаются при температуре 10—16 °С мороза, то осенью они могут пострадать и при менее низкой температуре. Например, корни сеянцев яблони лесной и груши лесной осенью начинают повреждаться при минус 3,5 °С. С конца сентября — начала октября устойчивость корней начинает повышаться и достигает максимума в декабре — январе, а в конце марта — апреле она резко снижается. Осенью корни часто повреждаются при запоздалой выкопке или же во время осенней транспортировки при снижении температуры окружающего воздуха до минус 3,5—6 °С и ниже. Осенью и в конце зимы наименее устойчивыми тканями корней являются кора, камбий и клетки камбиальной зоны; в зимнее время эти ткани более устойчивы, чем клетки первичной и вторичной ксилемы. В начале октября клетки камбия и вторичной флоэмы корней большинства плодовых пород повреждаются при температуре минус 3,5°, а в конце октября — начале ноября — при минус 5,7—6,2 °С. При повреждении коры и камбия корней раннеосенними заморозками на поверхности коры появляется плесень. Растения, на коре корней которых появилась плесень, непригодны для посадки. Плесень — признак гибели корней. В зимнее время гибель камбия, клеток камбиальной зоны и вторичной флоэмы тканей у корней яблони происходит при температуре от 10 до 12° мороза, а у более устойчивых форм — при минус 16 °С. В конце февраля — марте корни теряют и без того невысокую морозоустойчивость и повреждаются при температуре 7,5—8 °С мороза. В конце зимы, как и осенью, у корней повреждаются прежде всего камбий, клетки камбиальной зоны и вторичная флоэма, клетки ксилемы остаются неповрежденными. Устойчивость клеток первичной и вторичной ксилемы корней плодовых растений к низким температурам более стабильна. Они подвержены значительно меньшим изменениям под действием мороза, чем клетки камбия и флоэмы. Древесина повреждается обычно при температуре минус 10—12 °С. Как уже указывалось, характер повреждения корневой системы в разные годы неодинаковый. У сеянцев, саженцев и взрослых деревьев может быть повреждена древесина корней при полной сохранности коры, камбия н клеток камбиальной зоны. Чем тоньше пробковый слой на корнях, тем сильнее они повреждаются. Когда повреждена древесина корней, а кора и камбий здоровы, листья распускаются позднее, чем обычно, рост деревьев ослаблен. В первый год после повреждения листья имеют светло-зеленую окраску, часто меньших размеров. У плодоносящих деревьев осыпаются плоды. При сильном повреждении древесины корней растения плохо восстанавливаются. У молодых и взрослых плодоносящих деревьев, а также у сеянцев и саженцев наблюдается повреждение морозами древесины корней у корневой шейки и ниже при сохранности коры и клеток камбиальной зоны. При таком типе повреждения растения могут восстанавливаться при выращивании их в условиях оптимального увлажнения. Встречается иной тип повреждения — у корневой шейки корни здоровые, а глубже, на расстоянии 7 —11 см от нее, отдельными локализованными участками повреждены кора и камбий корней. На поврежденных участках коры появляется плесень. Еще глубже, на расстоянии 11 —17 см от корневой шейки, корни здоровые или имеют незначительное повреждение древесины. Мелкие мочковатые корни в верхних слоях почвы полностью погибли. При таком типе повреждения сеянцы и саженцы не восстанавливаются. При сильном повреждении коры и камбия корневой системы у молодых деревьев могут распуститься почки, образоваться листья, могут даже зацвести, а затем растения засыхают. Повреждение коры и камбия как у молодых, так и у взрослых растений — наиболее опасный тип повреждений корней. При таком типе повреждения кора у деревьев яблони отстает от древесины и снимается хлопьями, на коре ствола появляются ожоги коричневато-оранжевого цвета, у сеянцев и саженцев в зимней прикопке кора с корней сползает чулком, оголяя древесину. Растения с таким типом повреждения погибают в первый год после суровой зимы: весной или в середине лета независимо от влажности почвы и питания. У сохранившихся сильно поврежденных деревьев в последующие годы уменьшается прирост. Когда же часть основных корней вымерзла полностью, прирост растений очень слабый или совсем отсутствует. В случае сильного повреждения вегетативно размноженных подвоев яблони и айвы растения могут восстанавливаться за счет образования новых корней вблизи корневой шейки. При выращивании растений с частично поврежденной корневой системой в условиях постоянного равномерного оптимального увлажнения, когда вымерзают отдельные более тонкие корни, а ниже корневой шейки сохранились неповрежденными участки коры и часть корней разного диаметра, то на неповрежденных участках происходит интенсивное образование новых корней — растения хорошо восстанавливаются; при выращивании в условиях ограниченного или неравномерного увлажнения растения растут слабо и в последующие годы частично или полностью засыхают. По интенснвности окраски листьев в первый год после суровой зимы можно безошибочно определить степень повреждения корневой системы: при слабом повреждении коры и камбия и значительном повреждении древесины листья светло-зеленые, при сильном повреждении коры корней они мелкие, по краям коричневые, побеги тонкие. Пожелтение листьев при повреждении корней морозами, как и при засухе или избыточном увлажнении, обусловлено подавлением синтеза кинетина или аналогичных ему соединений, которые синтезируются в корнях растений и оттуда транспортируются в листья и молодые побеги, активизируя синтез белка. При угнетении роста корней, вызываемого иными факторами, листья обычно начинают желтеть вследствие распада белка и связанного с этим выцветания хлорофилла. С увеличением степени повреждения корневой системы молодых и плодоносящих деревьев прирост их уменьшается, листья образуются в нижней части дерева, на стволе вместо побегов часто появляются мелкие листья с явными признаками хлороза, типа розеточных. Во второй половине лета они опадают, на деревьях остаются крупные плодовые почки. Верхние побеги и ветви засыхают, а отдельные скелетные ветки и ветки первого порядка отмирают, деревья начинают суховершинить, а затем погибают. При сильном повреждении корневой системы плодоносящих деревьев, где камбий и клетки камбиальной зоны вымерзли, растения погибают. Если хотя бы на отдельных участках корней кора и камбий остались здоровыми, образуются новые корни, растения не погибают, но растут слабо. В тех случаях, когда повреждены концы корней второго порядка, прирост растений бывает слабым, листья становятся красновато-желтыми и осыпаются задолго до конца вегетации, в конце сентября — начале октября у них изменяется окраска коры, побегов и ветвей, кора приобретает светло-коричневый цвет. Характер проявления повреждения корневой системы у груши иной, чем у яблони. На мертвых корнях груши через год после повреждения морозами внешние признаки повреждения корней отсутствуют: кора на них гладкая, плотно прилегает к древесине. Повреждение можно обнаружить, только сделав поперечные и продольные срезы корней. Поврежденные ткани на срезах имеют темно-коричневую окраску. Неодинаковой морозоустойчивостью характеризуются корни различных плодовых пород. Из семечковых наименее морозоустойчивы корни айвы, груши лесной и некоторых форм яблони лесной, наиболее морозоустойчивы сеянцы яблони китайки, некоторые сорта яблони летнего и осеннего сроков созревания (Боровинка, Антоновка обыкновенная). Из косточковых наиболее морозоустойчивы сеянцы антипки и абрикоса. Саженцы сливы, абрикоса и даже персика, привитые на сеянцах абрикоса, сохраняются неповрежденными в бесснежные суровые зимы, а привитые на алыче — вымерзают. Корни антипки не имеют признаков повреждения при температуре почвы минус 22—23 °С на глубине узла кущения злаковых культур,минус 15,5° на глубине 20 и минус 12,8° на глубине 40 см, в то время как корни большинства саженцев плодовых пород сильно повреждаются при этих и даже более высоких температурах. Корни антипки более морозоустойчивы, чем корни яблони лесной и сеянцев культурных сортов. Наиболее морозоустойчивые формы китайки приближаются по степени устойчивости к антипке. Критическими температурами в зимнее время для корней большинства подвоев молодых растений яблони являются температуры 10—12 °С ниже нуля. Некоторые наиболее устойчивые формы могут выдерживать морозы до 16 °С и ниже, груши и черешни 9— 10, айвы — 8—9, персика — 10—11, абрикоса— 12—14, антипки — 15—16, земляники — 13—16 °С. Корневая система смородины и крыжовника может перенести не повреждаясь, температуру до минус 18 °С. Зная критическую температуру гибели корневой системы, температуру и глубину промерзания почвы, в зимнее время можно прогнозировать размеры повреждения корневой системы в том или ином районе и своевременно провести мероприятия по оздоровлению поврежденных морозом насаждений. Характер повреждения корней зависит от скорости снижения температуры и времени воздействия ее на корневую систему. При резком снижении температуры наблюдается сильное повреждение прежде всего коры и камбия даже в тот период, когда клетки этих тканей более устойчивы, чем древесина. Сердцевина у корней, кроме вегетативно размножаемых подвоев, обычно отсутствует. Степень повреждения корней зависит от наличия снежного покрова, его высоты, типа почвы, системы ухода за ней в саду и т. д. На легких супесчаных почвах корни плодовых деревьев повреждаются сильнее, чем на более плодородных, богатых органическими веществами. Песчаные почвы имеют большую теплопроводность, быстрее и на большую глубину промерзают, чем суглинистые; влажные промерзают медленнее, чем сухие. После засушливых лета и осени повреждение плодовых деревьев, особенно их корней, усиливается. В районах недостаточного увлажнения политые деревья лучше зимуют и меньше повреждаются морозами. В зависимости от системы содержания почвы в саду глубина и температура ее промерзания неодинаковы. Хорошо разрыхленная почва промерзает медленнее и на меньшую глубину, чем уплотненная. Мульчирование почвы (торфом, перепревшей соломой, перегноем) и окучивание деревьев хорошо предохраняют корневую систему растений от вымерзания. Внесение высоких норм органических удобрений, а также избыточное увлажнение приводят к повреждению и гибели корневой системы. При внесении высоких норм органических удобрений гибнут прежде всего кора, камбий и клетки камбиальной зоны не только обрастающих, но и скелетных корней, затем гибнет все растение. При избыточной влажности почвы вследствие ухудшения аэрации и повреждения корневой системы подавляются ростовые процессы и наблюдается ограниченное поступление воды и минеральных веществ в растение, поглотительная способность корней снижается в 2—3 раза по сравнению с корневыми системами деревьев, растущих в условиях оптимального увлажнения. При повреждении корневой системы в одних случаях происходит отмирание только всасывающих корней или же понижается их поглотительная способность, в других — отмирают всасывающие корни и повреждаются па-ренхимные клетки проводящих тканей (часто в клетках последних образуется вещество типа индиго, имеющее сине-фиолетовую окраску), в третьих — происходит гибель корней и всего растения. При избыточной влажности восстановительная активность тканей снижается, происходит смещение рН клеточного сока в кислую сторону, ухудшается состояние биоколлоидов протоплазмы. Изменяется пигментация листьев, на них появляются некрозы. При ограниченном и переменном увлажнении угнетаются ростовые процессы, подавляется образование поглощающих и развитие проводящих корней. На степень повреждения корневой системы плодоносящих деревьев оказывает влияние привой и прежде всего его физиологическое состояние: при повреждении штамба морозобоинами и солнечными ожогами морозоустойчивость корневой системы резко снижается. Более низкая ее морозоустойчивость отмечается также при несовместимости привоя с подвоем. Обильный урожай в год, предшествующий суровой зиме, снижает морозоустойчивость корневой системы. У молодых деревьев она отличается повышенной регенерационной способностью, у плодоносящих — пониженной. Восстановление плодоносящих деревьев возможно в тех случаях, когда повреждены только мелкие, мочковатые корни в верхних слоях почвы, а проводящие здоровые или же у них повреждена древесина, а камбий, клетки камбиальной зоны и кора здоровые или повреждены частично. При сильном повреждении камбия, клеток камбиальной зоны и коры деревья погибают в первый или последующий после суровой зимы год. ПОВРЕЖДЕНИЕ МОРОЗАМИ НАДЗЕМНОЙ ЧАСТИ Надземная часть плодовых деревьев в садах и саженцев в питомниках повреждается морозами не только в северных, но и в южных районах страны. Степень и характер повреждения весьма разнообразны и изменяются в зависимости от условий выращивания и сочетания метеорологических факторов. Характер повреждения зависит также от биологических особенностей сорта, возраста растений и продолжительности воздействия низкой температуры. Наиболее часто повреждаются морозами саженцы в питомниках, а также молодые деревья в садах в первые годы после посадки. Меньше повреждаются деревья, вступившие в пору плодоношения, в возрасте 10—12 лет, сильнее — взрослые плодоносящие деревья. Однако бывают случаи, когда деревья в 7— 13-летнем возрасте вымерзают, а 3—5- и 40-летнем сохраняются, как это было отмечено в зиму 1971/72 г. в садах колхозов и совхозов Винницкой, Хмельницкой и других областей Украины. Яблони сортов Пармен зимний золотой, Джонатан, Кальвиль снежный, Ренет Симиренко, Ренет ландсбергский в возрасте 7 —13 лет дали обильный урожай в год, предшествующий суровой зиме, сильно повредились и в последующие годы резко снизили урожайность. Часть этих деревьев погибла, в то время как более молодые и 40-летние деревья остались неповрежденными или были повреждены в слабой степени. Характер и степень повреждения надземной части разных плодовых пород в одну и ту же зиму и в разные зимы неодинаковы. По степени морозоустойчивости породы располагаются следующим образом: из семечковых — яблоня, груша, айва; из косточковых — вишня, слива, черешня, абрикос, персик. Такой порядок в устойчивости отдельных пород не всегда сохраняется. В зависимости от сочетания метеорологических факторов лета, осени и зимы, условий выращивания и физиологического состояния растений порядок расположения пород в отношении их устойчивости к низким и переменным температурам может быть иным. Так, в зимы 1965/66, 1971/72 гг. сильно повредились деревья яблони осенне-зимних сортов, часть из которых вымерзла полностью, сохранились неповрежденными или были повреждены в слабой степени деревья груши и сливы. Более сильная степень повреждения деревьев яблони обусловлена продолжительной деятельностью камбия: у яблони активная деятельность камбия заканчивается на 14—36 дней позже, чем у груши. В зимы, когда состояние камбия в осенний период предопределяет исход перезимовки плодовых деревьев, менее морозоустойчивые породы, характеризующиеся более короткой продолжительностью активной деятельности камбия, повреждаются слабее, чем более морозоустойчивые в целом породы, но отличающиеся более продолжительной деятельностью камбия. В суровые зимы нередки случаи вымерзания деревьев косточковых пород до линии снежного покрова. Наиболее часто у них вымерзают плодовые почки, плодовые веточки и концы однолетних побегов. Верхняя часть побегов и ткани,чрасположенные под почками (листовые следы) повреждаются сильнее, чем другие части побега. У косточковых пород, особенно у абрикоса, наблюдается кольцевое повреждение коры вокруг корневой шейки и в средней части ствола. У семечковых пород повреждаются не закончившие рост побеги, плодовые сумки, стволы и развилки. Наименее морозоустойчивыми являются побеги и плодовые веточки с хорошо развитой сердцевиной. Клетки сердцевины характеризуются низким содержанием углеводов, белков, фенольных соединений и минеральных веществ. В процессе роста побегов у отдельных пород она часто разрушается. Развитие сердцевины в побегах может служить одним из морфологических признаков степени морозоустойчивости растений или отдельных органов. МОРОЗОУСТОЙЧИВОСТЬ РАЗЛИЧНЫХ ТКАНЕЙ СТВОЛА, ВЕТВЕЙ И КОРНЕЙ Ствол и корень формируют целостную структуру растений. По анатомическому строению, характеру роста и выполняемым функциям они имеют общие особенности и различия. Корни большинства сеянцев подвоев не имеют сердцевины, а кора их сбрасывается в ходе вторичного роста. Сердцевина четко обнаруживается лишь у корней айвы и некоторых вегетативно размножаемых подвоев. В побегах, ветвях, стволах и плодовых образованиях сердцевина хорошо развита. В зимнее время она является наиболее чувствительной к морозам. Однако повреждение и даже полная гибель сердцевины не имеет значения для последующих процессов жизнедеятельности плодовых деревьев. Во внутренних клетках сердцевины протопласт отсутствует, многие из них отмирают к концу лета. Периферические клетки сердцевины, граничащие с ксилемой (перимедулярной зоной), живые. В них откладывается крахмал, редко используемый растением. Степень морозоустойчивости надземной части растения, как и корневой системы, подвержена сезонным изменениям. Диапазон температур, при котором происходят изменения, у надземной части значительно шире, чем у корневой системы. Он находится в пределах от 0,5—1,3 °С в период весенних заморозков до 42 °С и ниже зимой. С осени степень морозоустойчивости плодовых деревьев повышается, а к концу зимы снижается. Критические температуры гибели разных пород в зимнее время колеблются от минус 22—24 до минус 42 °С и ниже. Наиболее устойчивые сорта яблони в период глубокого покоя при хорошей подготовке их к зиме повреждаются при температуре минус 42—48 °С. При раннеосенних морозах или резких колебаниях температуры в середине зимы морозоустойчивость деревьев резко понижается даже у наиболее устойчивых сортов; у районированных по Украинской ССР сортов груши, сливы и вишни цветковые почки вымерзают при температуре минус 32—36 °С, в отдельные годы они повреждаются при температуре минус 26—30, черешни — минус 26—32 (или при минус 24—28 °С), абрикоса минус 24— 28 (после сухого лета или после поражения деревьев Monilia laxa минус 23—24), персика — минус 22—26 °С. При неблагоприятных условиях подготовки растений к зиме критические температуры гибели могут быть более высокими. Эти данные дают возможность определить границы распространения той или иной породы при сопоставлении абсолютных минимальных и критических температур гибели деревьев. При резких колебаниях температуры во второй половине зимы или же при внезапных морозах осенью плодовые деревья могут повреждаться и даже гибнуть при температуре 16—25 °С ниже нуля.
Кроме хорошо заметных, часто бывают повреждения, которые можно обнаружить только на поперечных и продольных срезах. К ним относится повреждение клеток флоэмы, камбия, камбиальной зоны, древесины и сердцевины ветвей, ствола, корней. Поврежденные морозом ткани обычно имеют бурую или коричневую окраску. Изменяется окраска их не сразу после воздействия низкой температуры, а спустя 20—30 дней осенью или в начале зимы и 5—10 дней в начале весны и в конце зимы. При воздействии низких температур на растущие части (во время заморозков или при замораживании в холодильной камере) повреждение может быть обнаружено через несколько часов. Неповрежденные сердцевина, древесина и камбий сохраняют белый или светло-серый цвет, а кора — зеленоватый. По интенсивности побурения и размеру повреждения ткани определяют степень повреждения. Ткани буреют также при поражении растений болезнями и повреждении вредителями, однако эти повреждения в большинстве случаев можно отличить от морозных. Поврежденные морозом клетки и ткани изменяют свои функции и структуру. Анатомические и гистохимические исследования показывают, что в побуревшей древесине трахеи и трахеиды полностью или частично заполняются веществами бурого цвета типа пектиновых камедей. Образование их происходит в основном за счет крахмала живых клеток, из которых они впоследствии попадают в сосуды. При превращении крахмала в камедь дерево лишается значительной части запасных питательных веществ. Закупорка сосудов камедью затрудняет продвижение питательных веществ и воды. Деревья яблони погибают, если у них закупорено больше 50 % сосудов. Для нормальной жизнедеятельности плодовых растений необходимо, чтобы большая 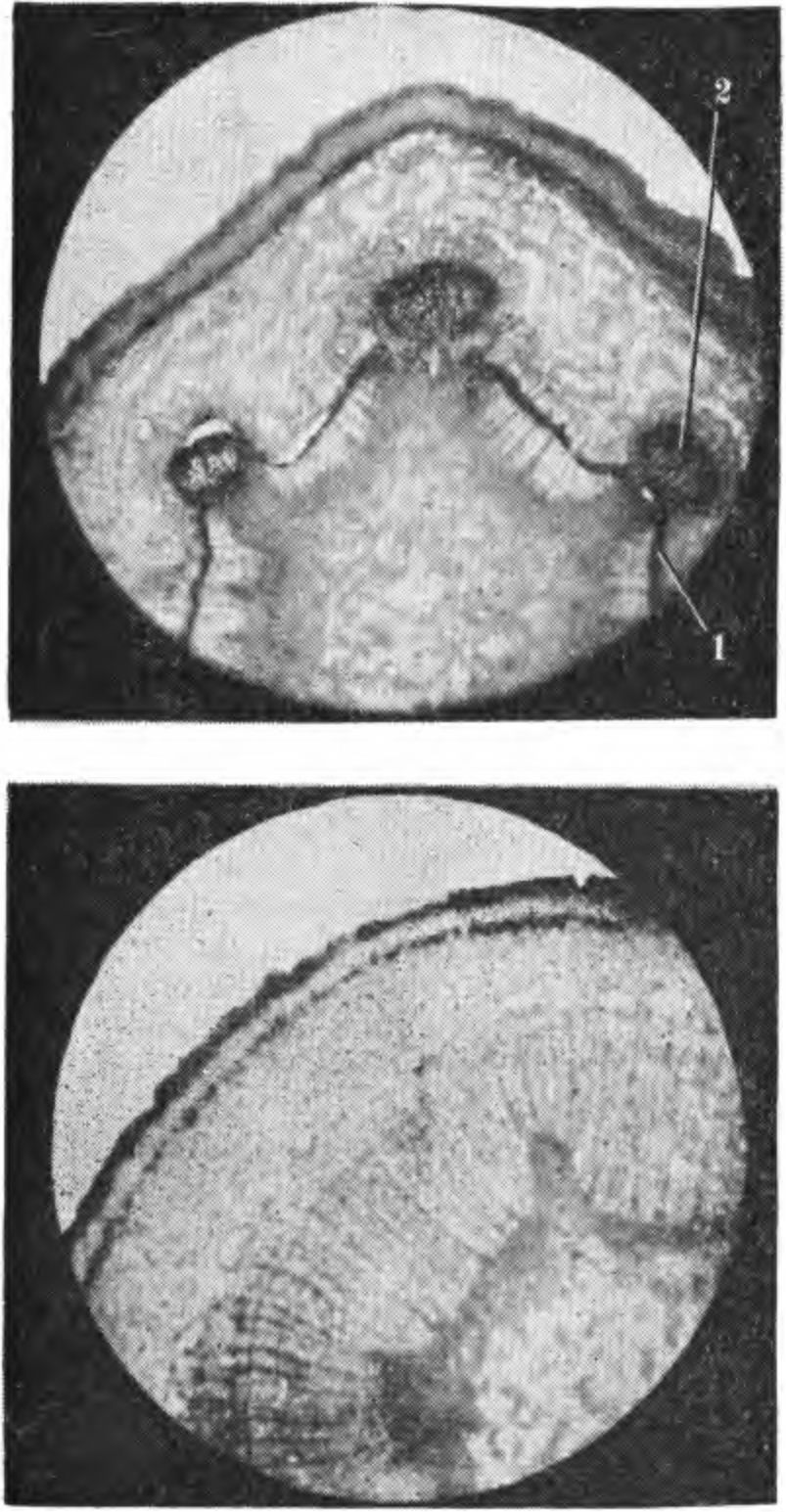
2. Повреждение побега яблони сорта Антоновка обыкновенная при замораживании: вверху — верхняя часть побега, повреждены камбий (1) и листовые следы (2); внизу — средняя часть побега повреждений не имеет. часть сосудов оставалась незакупоренной до тех пор, пока камбием не будет образован новый слой древесины (Steinmetz F. H.,Hil-born М. Т., 1937). Стенки клеток поврежденных морозом тканей подвергаются изменениям, резко снижающим механическую прочность древесины. Побурение древесины груши при зимних повреждениях, как известно, вызывается не образованием камеди, а иными превращениями углеводов и образованием дубильных веществ. Поврежденные морозом клетки коры, камбиальной зоны и древесины побегов и ветвей в процессе роста светлеют. Хорошо заметное весной или в начале лета побурение тканей к концу лета довольно часто исчезает, наблюдается так называемая резорбция. При сильном повреждении древесины у груши потемнение тканей сохраняется. Ветви и стволы с сильно подмерзшей древесиной становятся хрупкими, теряют прочность, отламываются отдельные скелетные ветви и даже разламываются стволы, на последних часто поселяются различные грибы и образуются дупла. На поперечных срезах ветвей отчетливо видны здоровые и поврежденные морозами годичные слои древесины. По этим слоям можно определить годы и размеры повреждений древесины морозами. Наиболее физиологически активным является самый молодой годичный слой древесины, прилегающий к камбию. С возрастом древесина теряет свои функции проводящей системы, клетки теряют воду и постепенно в них исчезает крахмал, ослабляется деятельность ферментов и нарушается контроль, регулирующий процесс жизнедеятельности клетки. Морозоустойчивость различных тканей надземных органов, как и корневой системы, непостоянна в разные периоды глубокого и вынужденного покоя. Наиболее подвержена сезонным изменениям морозоустойчивость камбия, клеток камбиальной зоны, коры вет вей и ствола, менее изменчивы клетки древесины (рис. 2—6). Клетки древесины образуют сложную в структурном и функциональном отношениях ткань, состоящую из живых и мертвых клеток (трахеид и члеников сосудов) проводящих воду, в сердцевинных лучах и отдельных паренхимных клетках ксилемы откладываются запасные вещества. Клетки ксилемы характеризуются пониженной метаболической активностью по сравнению с клетками флоэмы и камбия. В них сосредоточены так называемые эргастические вещества — крахмал, гемицеллюлоза, пектиновые и другие вещества, характеризующиеся высоким содержанием воды и упорядоченным расположением ее молекул. У древесных плодовых растений процесс крах-малообразования имеет сезонные различия и связан с активностью протопластов, гидролитических ферментов и физиологическим состоянием растений. Содержание крахмала и других углеводов существенно изменяется в разные периоды вегетации и покоя — увеличивается в коре ветвей и ствола осенью, а зимой при снижении температуры и усиливающихся гидролитических процессах в коре ветвей зимостойких сортов крахмал исчезает быстрее, чем у слабозимостойких, затем вновь появляется весной. При сильном повреждении древесины ветвей и ствола морозами у растений ослабляются ростовые процесы, резко снижается последующая зимостойкость и продуктивность, на коре появляется сажистый налет. Наличие такого налета является хорошим морфологическим признаком для определения подмерзания древесины у деревьев яблони и груши (Соловьева М. А., 1958). Повреждение древесины ствола и ветвей морозом определить сразу трудно. В зависимости от водного режима и типа почвы, уровня минерального питания, рельефа местности и биологических особенностей сорта реакция на подмерзание проявляется в разной степени и не в одно и то же время. Нередко у деревьев с поврежденной древесиной в результате нарушения деятельности проводящей системы наблюдается мелколистность, хлороз и млечный блеск. В последующие суровые зимы такие деревья могут вымерзнуть. При повреждении надземной части растений обычно снижается зимостойкость корневой системы. Саженцы яблони, груши, сливы и вишни со слабо- и среднеповрежденной древесиной в условиях оптимального увлажнения могут нормально развиваться. При ограниченной же влажности почвы они растут плохо и в последующие зимы вымерзают. У саженцев с сильно подмерзшей древесиной в нижней части ствола пробуждаются спящие почки и образуются побеги, которые усиленно ветвятся, приобретая вид куста. Такие растения теряют производственную ценность. У некоторых деревьев в саду, а также у саженцев, древесина ствола которых была повреждена морозом в сильной степени, на второй и третий годы после суровой зимы начинают появляться ожоги и морозобоины. При повреждении древесины скелетных ветвей и ствола процессы регенерации у разных сортов происходят неодинаково. Не у всех сортов размер поврежденной ткани остается таким же, как и в год повреждения. У одних зона повреждения древесины расширяется (Ренет Симиренко, Кальвиль снежный, Бойкен, Пармен зимний золотой, Ренет ланд-сбергский, Августовское и др.), у других она как бы локализуется, остается такой же, какой была в год повреждения (Мекинтош, Пепинка литовская и др.), т. е. образуется изолирующий слой, предохраняющий от распространения повреждения. У деревьев, древесина которых повреждена в сильной степени, не только ухудшается водный режим, но и изменяется направленность деятельности ферментных систем, кроме того активизируется дыхание, особенно в зимнее время, и снижается последующая зимостойкость деревьев. Саженцы, у которых повреждена древесина (больше 50 % общей площади среза), а кора ствола имеет сажистый налет, непригодны для посадки в саду. Если такие растения повреждены в питомниках, их необходимо срезать на повторный рост, а растения более старшего возраста — выкорчевать, заменив здоровыми. Повреждение камбия надземных органов плодовых деревьев наблюдается часто. Оно приводит к структурным и функциональным изменениям физиологических процессов. При повреждении морозом камбия клетки вторичных ксилемы и флоэмы не образуются, ухудшается оводненность тканей, нарушается поступление органических и минеральных веществ в растение. Камбий обычно возникает из прокамбия и характеризуется сезонными морфологическими и функциональными изменениями. Относится к латеральным меристемам с тангентальным типом деления клеток, образующим вторичные проводящие ткани. Располагается между ксилемой и флоэмой и имеет два типа клеток: удлиненные с заостренными концами — веретеновидные (прозенхимные) инициал и и с несколько меньшими по размеру клетками лучевых инициалей, отличающихся своими функциями и строением. Веретеновидные инициали образуют элементы ксилемы и флоэмы, лучевые являются предшественниками образования клеток лу- 3. Повреждение побега яблони сорта Ренет ланде-бергский при замораживании: вверху — в период вынужденного покоя, повреждены сердцевина (1), частично древесина (2), камбий (3) и кора (4); внизу — в конце периода вынужденного покоя, погибли камбий (3) и кора (4). 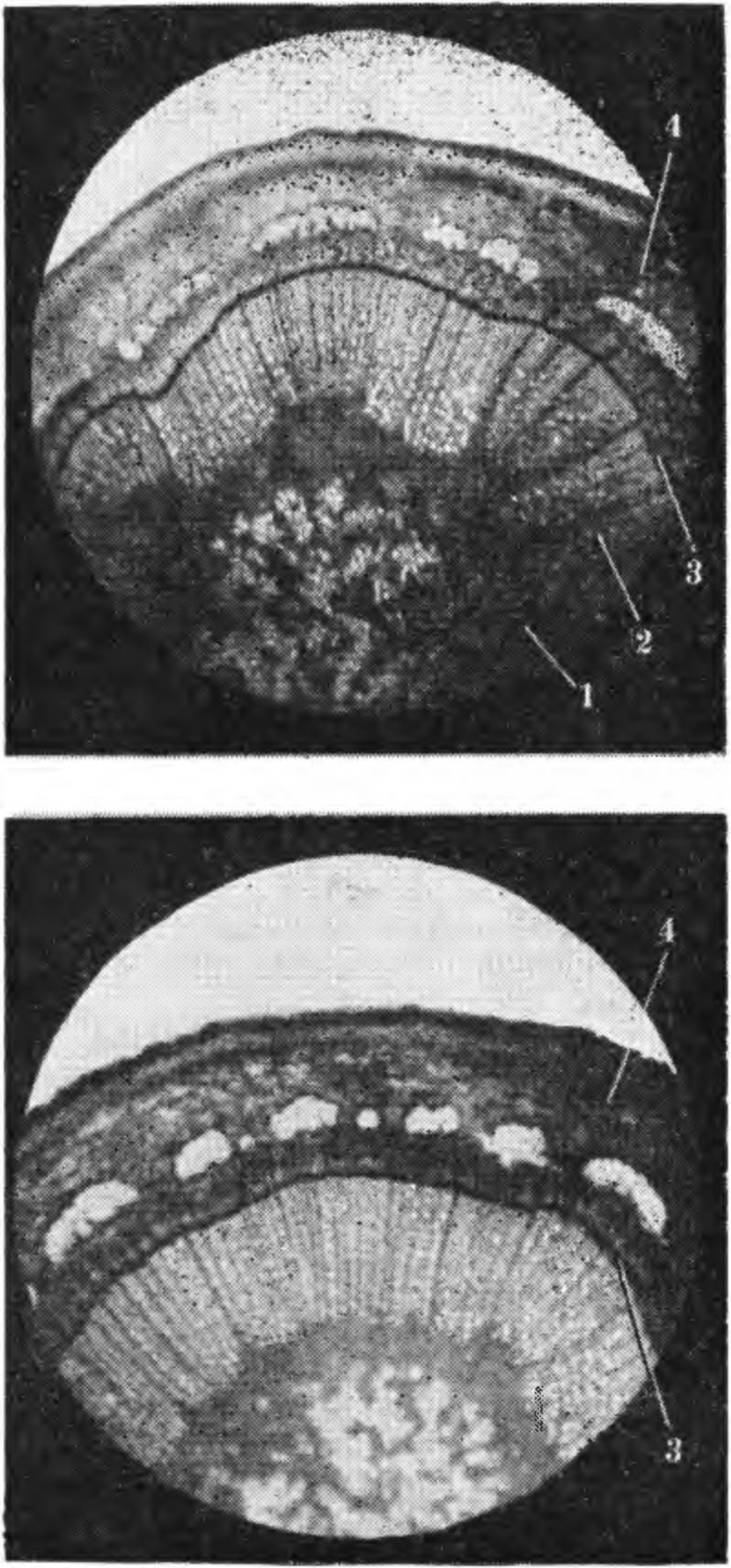 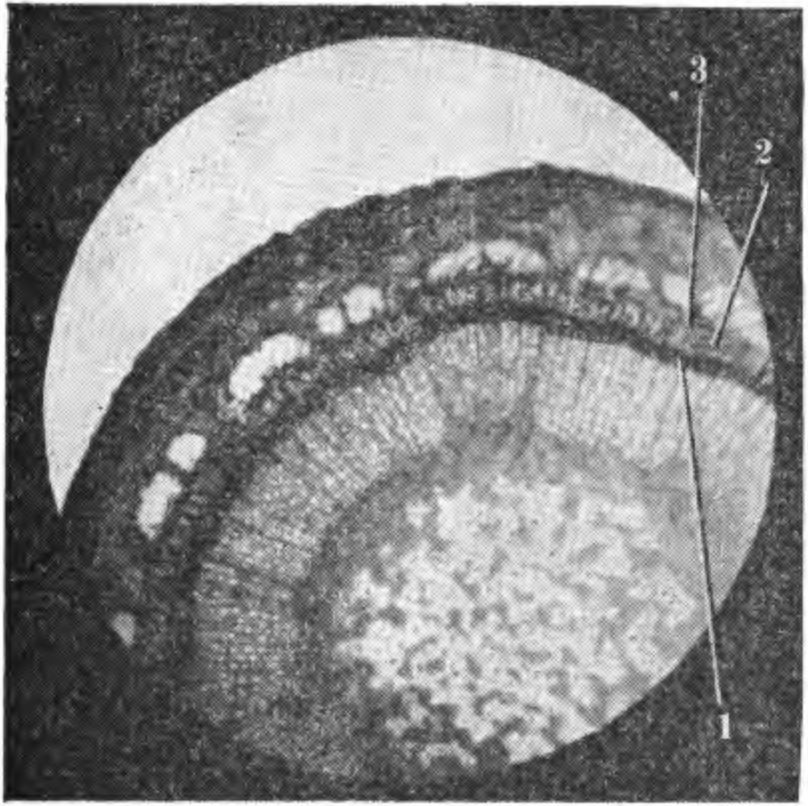
4. Повреждение наиболее молодого годичного кш1ь-ца (1) древесины у яблони сорта Ренет ландсберг-ский при замораживании; образование нового слоя древесины (2) произошло в летний период в результате деятельности камбия (3). чевой или поперечной системы ксилемы и флоэмы. В период активного роста камбиальные клетки сильно вакуолизируются, стенки клеток и слой протоплазмы утончаются, вязкость ее уменьшается, обнаруживается быстрое движение протоплазмы внутри клеток. В этот период камбий трудно различим — он сливается с клетками камбиальной зоны. Инициали и их производные образуют зону мерисгематических клеток, располагающихся в виде правильных радиальных рядов, называемых камбиальной зоной. Мелкие ^еристематические клетки камбиальной зоны увеличиваются в размере, дифференцируются с образованием ксилемы и флоэмы. Камбиальную активность предопределяет физиологическое состояние растений. В период активной деятельности камбия наблюдается легкое отделение коры от древесины. Если деление клеток и дифференциация тканей в камбильной зоне приостанавливается, кора перестает отделяться от древесины. В период покоя камбий легко можно обнаружить на границе между флоэмой и ксилемой. В этот период камбиальные клетки обычно заполнены протоплазмой с замедленным движением и повышенной вязкостью. В них появляются масла, исчезающие в период активной деятельности камбия. Установлена сезонная периодичность активной деятельности камбия. Начало ее в Лесостепи и на Полесье Украины отмечено в первой, иногда во второй декаде апреля. У различных пород и сортов плодовых деревьев существенных различий в начале активной деятельности камбия не имеется. При неблагоприятном сочетании метеорологических факторов и плохих условий выращивания активная деятельность камбия прерывается, а при нормализации условий вновь восстанавливается; древесина в такие годы образует два кольца, одно из которых ложное, или множественное. Годичные кольца обычно отчетливо видны на поперечных срезах ветвей и ствола. Активная деятельность камбия раньше заканчивается у однолетних ветвей и позже — у основания ствола и в развилках. Разница в окончании активной деятельности камбия в разных органах одного и того же растения составляет от 5—11 до 38 и более дней. Более короткой камбиальной деятельностью в пределах одной породы отличаются морозоустойчивые сорта. У зимостойких сортов яблони активная деятельность камбия в однолетних побегах заканчивается во второй половине августа, иногда в начале сентября, а у слабозимостойких — во второй половине сентября или во второй и даже третьей декаде октября. Разница в окончании этой деятельности у разных по зимостойкости сортов составляет 49—50 и даже 74—76 дней. У груши и вишен активная деятельность камбия часто заканчивается в более ранние сроки, чем у самых зимостойких сортов яблони. Следовательно, в годы, когда повреждение 5. Поврежденные при замораживании побега груши сердцевина (1), древесина (2) и листовые следы (3) (фото А. Д. Чижа). 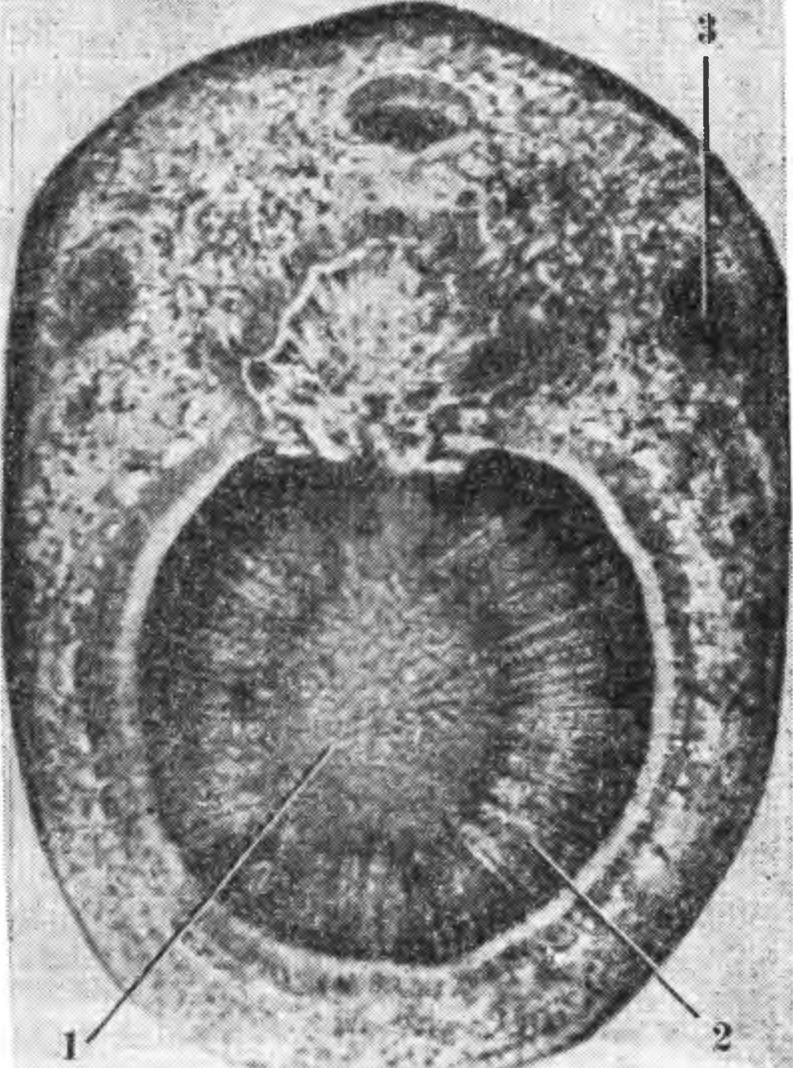
плодовых деревьев наблюдается при наступлении раннеосенних морозов и бывает обусловлено гибелью клеток камбия, деревья груши, отличающиеся намного меньшей морозоустойчивостью в зимнее время по сравнению с яблонями, повреждаются слабее или совсем не повреждаются. В этот период однолетние ветви более морозоустойчивы, чем 3—4-летние ветви и стволы, особенно у молодых деревьев. В различных частях побега активная деятельность камбия заканчивается неодновременно: в междоузлиях раньше, возле почек — позже. В конце зимы — начале весны у более морозоустойчивых сортов камбий часто начинает возобновлять активную деятельность раньше, чем у менее устойчивых к морозу. При резком снижении температуры в конце февраля — марте наблюдается повреждение камбия, клеток камбиальной зоны и коры побегов и ветвей у морозоустойчивых сортов, а у неморозоустойчивых они не повреждаются. На продолжительность активной деятельности камбия и степень его морозоустойчивости большое влияние оказывают водный режим, оводненность тканей и уровень обеспеченности растений минеральными веществами, особенно азотом, калием и кальцием. Калий и кальций способствуют сокращению активной деятельности камбия и снижению интенсивности дыхания зимой, оказывают влияние на углеводный метаболизм, способствуют повышению содержания запасных резервных углеводов и олигосахаридов. Марганец и бор как бы стабилизируют степень устойчивости этих тканей в конце зимы (Соловьева М. А., Оканенко А. С., 1970). Камбий и клетки камбиальной зоны отличаются высокой степенью метаболической активности. Камбий — одна из наиболее жизненно необходимых постоянно возобновляющихся тканей. При сильном повреждении клеток камбия стволов и развилок, угнетаются ростовые процессы, изменяется окраска и размеры листьев и плодов. Листья образуются мелкие, светло-зеленые, по краям усыхают. Засыхают также и недоразвитые плоды. К осени деревья с сильным повреждением камбия ствола и развилок засыхают. Если камбий ветвей сильно поврежден, сначала на коре появляются различные красновато-желтые пятна и трещины, которые, увеличиваясь в размере, приводят к гибели отдельных ветвей или всего дерева. При повреждении камбия ветвей кора у деревьев отмирает чаще на нижней стороне их, у саженцев — в средней части ствола. Устойчивость камбия и клеток камбиальной зоны в зависимости от уровня минерального питания, оводненности тканей, обрезки и других агротехнических приемов может существенно изменяться. Избыточное азотное удобрение, как и неравномерное увлажнение 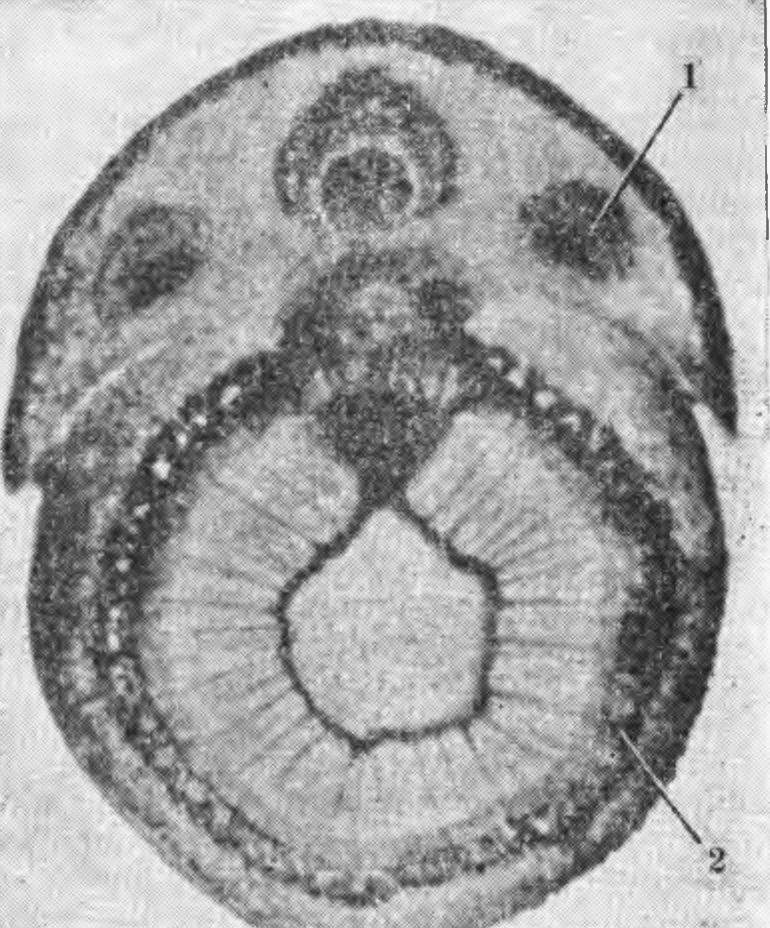 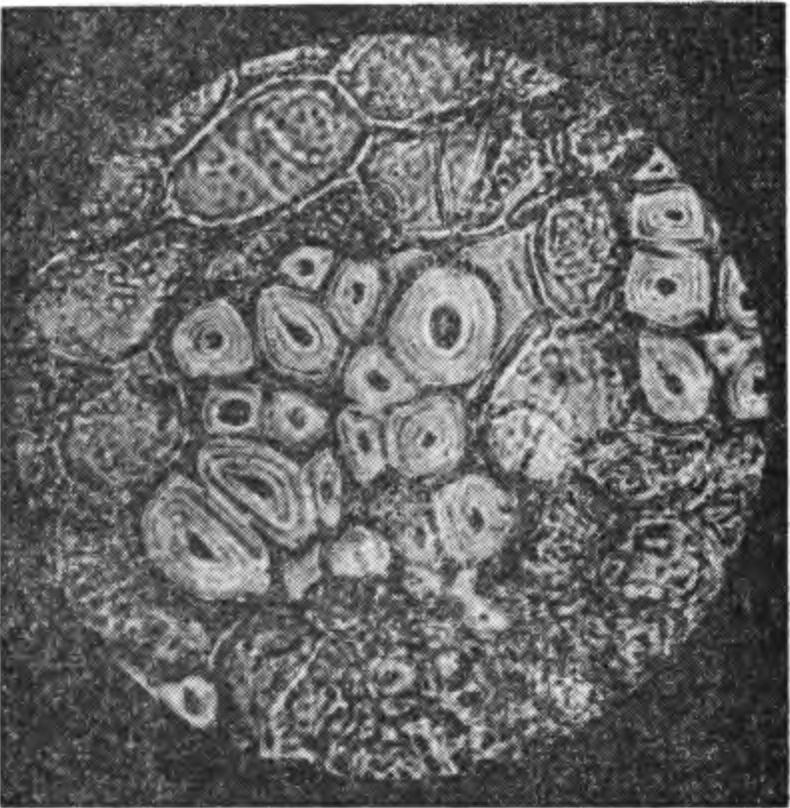
6. Вверху — повреждение листовых следов (1) и паренхимных клеток флоэмы (2), окружающих склеренхиму. Внизу — клетки склеренхимы в увеличенном размере (фото А. Д. Чижа). или переувлажнение почвы, резко снижают устойчивость камбия к низким и переменным температурам в осенне-зимний и зимне-весенний периоды. Сильная обрезка весной или обрезка недостаточно устойчивых сортов в зимнее время приводит к затяжному росту и обусловливает снижение морозоустойчивости камбия и клеток камбиальной зоны, вследствие чего увеличивается возможность повреждения этих тканей морозами в разные периоды осени, зимы и весны. Окончание активной деятельности камбия сопровождается усилением биосинтеза антоцианов. ПОВРЕЖДЕНИЕ КОРЫ СТВОЛА И СКЕЛЕТНЫХ ВЕТВЕЙ Кора является компонентом гетерогенных тканей и выполняет функции проводящей системы. Паренхимные клетки ее синтезируют и накопляют различные вещества. Они образуют радиальные лучи и вертикальные ряды живых клеток во флоэме. Повреждение коры плодовых деревьев морозами имеет разнообразный характер, проявляется в виде небольших, вначале малозаметных пятен или трещин. Часто повреждаются не только наружные слои клеток коры, но и ее внутренние элементы, в результате чего происходит необратимая деструкция пластид, их агглютинация и другие сложные структурные изменения протоплазмы. Под влиянием низких критических температур в зимний период разрушается структура хлоропластов, набухают стромы и одновременно разрываются их оболочки. Наблюдается изменение свойств цитоплазмы. Нарушение проницаемости ее тонопластной мембраны вызывает диффузию вакуолярного содержимого по всему объему клетки. Данное явление, как отмечает А. В. Брайон (1973), необратимо и приводит или к отмиранию отдельных участков коры ветвей либо к полному повреждению хлорофиллоносных клеток многолетних ветвей и ствола плодовых растений. Осенью и ранней весной, когда при резких колебаниях температуры повреждаются камбий и клетки камбиальной зоны, клетки вторичной флоэмы и паренхимные клетки коры, окружающие склеренхиму, клетки ксилемы, сохраняются неповрежденными. В зимнее время клетки первичной и вторичной флоэмы более устойчивы, чем клетки ксилемы. Но не все клетки фюэмы одинаково устойчивы. Наименее устойчивы клетки, окружающие склеренхиму, наиболее устойчивы клетки вторичной флоэмы и камбиальной зоны. Склеренхимные клетки встречаются как в виде тканевых компонентов, так и небольшими группами. Это непаренхимные клетки, имеющие многогранную форму. У груши и яблони они выполняют механическую функцию. Склеренхимные клетки имеют толстые вторичные оболочки, расположенные в виде прерывающейся цепочки в зоне клеток вторичной флоэмы. Клетки склеренхимы лишены протопластов, в связи с чем и функциональная деятельность их ограничена. В коре при морозных повреждениях иногда наблюдается процесс преобразования па-ренхимных клеток в эмбриональные с последующей их дифференциацией. Клетки флоэмы отличаются более высоким уровнем метаболической активности и функциональной специализации, чем клетки ксилемы. Они характеризуются более высоким содержанием органических и минеральных веществ. Это сложная в физиологическом значении ткань, выполняющая разно образные функции. Паренхимные клетки флоэмы яблони, абрикоса и других плодовых культур со второй половины лета, в период подготовки растений к зиме, накапливают крахмал, гемицеллюлозу, жиры, фенольные соединения и особенно антоцианы, которым принадлежат защитные функции, связанные с изменением фосфорного метаболизма и степенью подготовки растений к зиме. Изменения фосфорного метаболизма сопровождаются повышением содержания фосфора эфиросахаров и неорганического фосфора. Последний принимает участие в осмотических явлениях клетки, а эфиросахара влияют на изменение состояния воды в ней и стабилизацию клеточных структур Соловье ва М. А., 1970). Повреждения морозами клеток первичной и вторичной флоэмы и других тканевых элементов коры разнообразны по степени и характеру, соответственно и повреждающий эффект их различен. При повреждении отдельных участков коры, как и древесины, образуется некротический (изолирующий) слой из продуктов распада поврежденных паренхимных клеток. В процессе роста изолирующий слой иногда разрушается. Как известно, при частичном повреждении ситовидных трубок на оболочках живых клеток откладывается каллоза и другие полимерные вещества, образующие прослойку между поврежденными и здоровыми тканевыми элементами. Эта прослойка преграждает дальнейшее распространение повреждения и отмирания клеток. При сильном повреждении клеток флоэмы перестает функционировать проводящая ткань, которая постепенно разрушается и отмирает. В зависимости от степени и характера повреждения отмирают отдельные поверхностные участки коры или вся кора до камбия. При сильном повреждении коры ствола или ветвей деревья плохо восстанавливаются. Деревья, имеющие кольцевое повреждение коры у корневой шейки или в средней части ствола, совсем не восстанавливаются. При полной гибели камбия, когда кора отстает от древесины и затем загнивает или засыхает, деревья также не восстанавливаются. Повреждение коры, как и камбия, зависит от сочетания метеорологических факторов лета, осени и зимы, а также от условий выращивания, обусловливающих физиологическое состояние растения. Наиболее опасный тип повреждения — повреждение камбия, клеток камбиальной зоны и коры ствола. К широко распространенным и опасным типам повреждения относятся солнечные ожоги и морозобоины, которые наблюдаются не только в северных и северо-восточных, но и в южных районах нашей страны. Исключением являются районы с повышенной относительной влажностью воздуха. ПОВРЕЖДЕНИЕ КОРЫ РАЗВИЛОК И СТВОЛА В ЗАВИСИМОСТИ ОТ УСЛОВИЙ ВЫРАЩИВАНИЯ Типы повреждения коры развилок и ствола плодовых деревьев весьма разнообразны. На коре ветвей в развилках и на стволе при повреждении морозами появляются темные пятна различного размера и разной формы, часть пятен имеет гладкую поверхность, часть — с поверхностными или глубокими продольными трещинами, края которых часто рваные и закручиваются наружу, часть — с корой, как бы вдавленной во внутрь. При сильном повреждении кора хлопьями сползает, оголяя древесину. В тех случаях, когда камбий под погибшей корой сохраняется живым, образуются новые клетки вторичных флоэмы и ксилемы. В дальнейшем происходит восстановление поврежденных морозом тканей. На деревьях многих осенне-зимних сортов яблони и груши, а также большинства сортов черешни, абрикоса и сливы наблюдается повреждение морозами коры в развилках ветвей, чаще встречающееся у молодых деревьев при вступлении их в пору плодоношения и в первые годы плодоношения, чем у взрослых плодоносящих деревьев. При повреждении развилок кора отмирает с внутренней стороны ветви. При резких колебаниях температуры часто повреждается кора в нижней ее части. Повреждения такого типа зависят от продолжительности ростовых процессов, прежде всего от окончания активной деятельности камбия, сочетания метеорологических факторов в послеморозный период и условий выращивания. Активная деятельность камбия в развилках заканчивается позже, чем на других участках ствола и ветвей, следовательно, подготовка этих участков коры к зиме происходит в более поздний период, отчего сдвигаются процессы метаболизма, оказывающие влияние на структурные изменения и энергетические процессы в клетках растений, что увеличивает вероятность повреждений этих тканей. Деятельность камбия и ее продолжительность, находясь в тесной связи с влажностью почвы, оводненностью тканей, относительной влажностью воздуха, наличием питательных веществ в почве, оказывает влияние на величину прироста древесины и морозоустойчивость растений. Наиболее отзывчивы плодовые деревья на азотные удобрения. При увеличении норм азота или при совместном внесении азота и фосфора в почву снижается морозоустойчивость растений. При внесении высоких норм азота наблюдаются изменения в фосфорном метаболизме, величине рН клеточного сока, содержании осмотически активных веществ, в клеточном соке уменьшается содержание Сахаров и крахмала (резервных углеводов, превращающихся при воздействии низких температур в процессе гидролиза в сахара и жиры, являющиеся защитными веществами), увеличивается продолжительность активной деятельности камбия, усиливаются ростовые процессы, которые до наступления заморозков не заканчиваются. Деревья яблони, сливы, груши, абрикоса, произрастающие при повышенном азотном питании или азотной недостаточности, повреждаются сильнее выращиваемых в оптимальных условиях питания. При азотной недостаточности листья растений приобретают желтоватый оттенок и преждевременно осыпаются, прежде всего желтеют листья розеток, а на листьях абрикоса и персика появляется дырчатая пятнистость. Эти растения слабо растут, кора у них становится желтовато-коричневой, восприимчивой к солнечным ожогам. Тип повреждения коры ствола при азотной недостаточности несколько отличается от повреждений, наблюдающихся при внесении повышенных доз азота. В последнем случае от мороза полностью погибают кора и камбий в развилках и в средней части ствола, кора приобретает темно-коричневую окраску с разными оттенками. Большинство таких деревьев погибает. При азотной недостаточности повреждаются поверхностные или более глубокие слои коры. В первом случае кора шелушится, при слабом повреждении растения оздоравливаются в процессе роста. При повреждении более глубоких слоев коры отмирают отдельные ее участки. Процессы оздоровления зависят от степени повреждения коры. Деревья, выращенные в условиях повышенного азотного питания, в суровые зимы сильно повреждаются и часто полностью вымерзают. Снижение зимостойкости при выращивании в таких условиях наблюдается не только у слабозимостойких сортов или пород, но и у наиболее зимостойких, таких как яблоня сорта Антоновка обыкновенная. Для условий Лесостепи и Полесья оптимальными являются следующие нормы удобрений, кг/га: азота 90, фосфора и калия по 120. Фосфорно-калийные удобрения, вносимые по фону азотных весной или в первой декаде августа, способствуют повышению морозоустойчивости коры, камбия и клеток камбиальной зоны. При внесении оптимальных норм азота, фосфора и калия плодовые деревья повреждаются морозами меньше, чем неудобренные или же выращиваемые при повышенном азотном питании. Они характеризуются лучшим ростом и более высокой урожайностью. Фосфорно-калийные удобрения способствуют сокращению камбиальной деятельности и повышению содержания олигосахаридов, часть из которых является ингибиторами ростовых процессов. Особая роль в зимостойкости плодовых растений принадлежит калию, принимающему участие в регуляции ростовых процессов, осмотических явлений, а также оказывающему положительное влияние на синтез полимерных соединений, фотосинтез и интенсивность дыхания в зимнее время. При азотной недостаточности внесение фосфорно-калийных удобрений снижает зимостойкость коры ствола и ветвей, ослабляет рост деревьев, листья становятся мелкие, светло-зеленые, как на деревьях, произрастающих на бедных супесчаных почвах без удобрений. При отсутствии азота наблюдается синергетическое действие отрицательного эффекта солей калия и фосфора, связанного с депрессией жизненных процессов и усилением напряженности фосфорного метаболизма. Под влиянием удобрений регулируется последовательность отдельных этапов в жизненном цикле растений, изменяется метаболическая активность тканей, продолжительность ростовых процессов, зимостойкость отдельных тканей и в целом плодового дерева. Более высокая зимостойкость плодовых деревьев наблюдается в тех случаях, когда весной вносят азотные удобрения, а во второй половине вегетационного периода дополнительно и фосфорно-калийные. Положительное влияние на зимостойкость оказывает также внесение весной фосфорно-калийных удобрений из расчета P90К120-135 по фону азота. Сильная обрезка снижает зимостойкость плодовых деревьев в северных, северо-восточных и западных районах Украинской ССР. После нее на коре ствола, развилок и скелетных ветвей образуются глубокие трещины или желтовато-коричневые полосы. С южной и юго-западной сторон вдоль всего ствола и ветвей первого яруса поврежденная кора отмирает, а затем отстает от древесины, последняя нередко трескается. Обычно деревья после сильной обрезки быстро погибают. При зимней обрезке яблони осенне-зимних сортов в северных, северо-восточных и западных районах Украины погибают ткани коры вокруг места среза ветви. Эти повреждения часто увеличиваются в размерах, у таких деревьев в дальнейшем резко снижается зимостойкость как надземной части, так и корневой системы. Подобный тип повреждения после зимней обрезки или сильной обрезки весной или в конце зимы, а также после поздней вырезки побегов утолщения на кольцо наблюдается во всех районах нашей республики. При перепрививке яблони, вступившей в пору плодоношения, морозоустойчивость коры ветвей часто снижается, особенно когда перепрививку всего дерева проводят в один год и удаляют большинство обрастающих ветвей. Такая перепрививка приводит к уменьшению листовой поверхности дерева, а следовательно, и к уменьшению содержания органических веществ — ассимилятов, вырабатываемых в процессе фотосинтеза и относящихся к защитным веществам. Ассими-ляты способствуют утолщению клеточных оболочек и ускоряют процесс лигнифика-цин. Лигнпфикацмя клеточных оболочек препятствует иссушающему действию межклеточного льда и защищает содержимое клеток от механического воздействия последнего. При перепрививке 6—10-летних деревьев не в один, а в два года и сохранении всех имеющихся обрастающих ветвей кора значительно меньше повреждается низкими отрицательными температурами возле мест среза ветвей. Уменьшается повреждение коры на ветвях также в тех случаях, когда прижившиеся черенки и хорошо развитые запасные ветви не вырезают на второй год после прививки, а пригибают, ослабляя их рост, а через 3—4 года вырезают. Особое значение в формировании морозо устойчивости и продуктивности плодовых деревьев имеет листовая поверхность, ориентация листьев в кроне и их физиологическое состояние. Повреждение листьев паршой, мучнистой росой и другими болезнями приводит к резкому снижению морозоустойчивости плодовых деревьев. В специальных опытах, проведенных нами в 1977 г. с удалением листьев в конце вегетации у деревьев яблони сортов Мекин-тош и Ренет Симиренко, было отмечено резкое снижение морозоустойчивости —20 % деревьев Мекинтоша полностью вымерзло в зиму 1977/78 г., а 80 % растений имели сильное повреждение коры у корневой шейки и в средней части ствола. Контрольные растения с хорошо сохранившимися листьями повреждений не имели. У поврежденных растений отмечено нарушение синхронизированной организации физиологических функций весной и летом. В течение следующих трех лет раны у корневой шейки поврежденных растений не заросли полностью. У этих растений ухудшался водообмен и снижалась водоудерживающая способность тканей в зимнее время, подавлялся синтез сложных углеводов и некоторых веществ фенольной природы — антоцианов и хал конов. В первой половине лета у поврежденных растений содержание Сахаров в коре ветвей уменьшалось в 5,2 раза, крахмала —2,8, гемицеллюлоз — 3,1, антоцианов —1,4 раза, халконов —3,3 раза, в древесине Сахаров —2,9, крахмала —2,5 и гемицеллюлоз в 4,7 раза. Накопление крахмала само по себе не повышает морозоустойчивости растений, но является необходимым условием для развития высокой морозоустойчивости. При гидролизе крахмала, гемицеллюлоз и других полисахаридов образуются защитные вещества, способствующие стабилизации гелизированных структур протоплазмы и перестройке ее мембранной системы. У поврежденных растений резко снижалась последующая морозоустойчивость отдельных тканей (камбия, клеток камбиальной зоны и вторичной флоэмы). Цветковые почки у этих растений не образовывались в течение двух последующих лет. Не только у яблони, но и у других плодовых растений, листья которых в течение лета поражались болезнями, вредителями или имели функциональные нарушения, резко снижалась морозоустойчивость цветковых почек, а также коры ствола, ветвей и корней. Это относится даже к таким морозоустойчивым растениям, как смородина черная. По данным 3. А. Шестопал (1969), при поражении растений антракнозом листья у смородины опадают на 1,5—2 месяца раньше естественного листопада, урожайность уменьшается в 3—5 раз и понижается зимостойкость. Гибель плодовых почек у этих растений в зимы с резкими колебаниями температуры достигает 100 %, надземная часть растений полностью вымерзает. Непораженные растения сохраняются и обильно плодоносят. Разной морозоустойчивостью характеризуются молодые деревья яблони с разной формой кроны. Деревья с плоской кроной, оптимальным радиационным режимом, исключающим эффекты взаимного затенения листьев, более устойчивы к низким и переменным температурам, чем с обычной округлой формой кроны. При формировании деревьев имеет значение степень ажурности кроны — естественной или искусственно создаваемой обрезкой. Загущение кроны приводит к снижению как общей продуктивности растений, так и их морозоустойчивости. Оптимизация радиационного режима предопределяет продуктивность и устойчивость плодовых растений к неблагоприятным факторам среды. С улучшением радиационного режима у плодовых деревьев повышается интенсивность фотосинтеза (у яблони в 1,5—2 раза), увеличивается содержание запасных защитных веществ, стабилизируется водоудерживающая способность тканей, повышается продуктивность и морозоустойчивость растений (Соловьева М. А., Починок X. Н., Оканенко А. С., 1965). Велика роль осеннего фотосинтеза в подготовке растений к зиме и их зимостойкости. Применение легкой омолаживающей обрезки оказывает влияние на биосинтез хлорофилла, содержание которого у омоложенных деревьев возрастает на 15.8—20,6 %. Увеличивается также содержание защитных веществ в тканях растений. У таких деревьев преждевременно не опадают листья, повышается интенсивность фотосинтеза. Так, в наших опытах весной у омоложенных деревьев Антоновки 22-летнего возраста она увеличивалась на 16,7 %, Кальвиля снежного — на 24,6 %, а во второй половине сентября у деревьев Антоновки до 28,2 %, Кальвиля снежного — до 10,3 % при средних значениях у контрольного дерева 4,80—4,92 мг С02 на дм 2/час (Соловьева М. А., Починок X. Н., Оканенко А. С., 1965). Урожай с одного омоложенного дерева Кальвиля снежного составлял 440 кг против 286 кг у неомоложенных контрольных деревьев. Урожайность повышалась, а морозоустойчивость деревьев не имела существенных различий. У нальметтных деревьев различно ориентированные в пространстве ветви характеризуются неодинаковой морозоустойчивостью. Наименее устойчивы центральные проводники деревьев с интенсивно развитой ксилемой. Морозоустойчивость наиболее метаболически активных тканей у ветвей, пригнутых вниз, в 2 раза, а растущих горизонтально в 1,8 раза выше, чем у центральных проводников. Низкая морозоустойчивость центрального проводника обусловлена замедленным синтезом запасных защитных веществ и локализацией ауксинов. Пониженной зимостойкостью характеризуются листовые следы, где расположена общая проводящая система. В годы с неблагоприятным сочетанием метеорологических факторов повреждаются прежде всего эти ткани, кора вблизи почек темнеет, растрескивается, как бы вдавливается во внутрь и отмирает. Влажность почвы оказывает существенное влияние на степень зимостойкости коры, камбия и клеток камбиальной зоны ствола, развилок и ветвей. Резко снижается зимостойкость коры и камбия надземной части деревьев при выращивании их в условиях избыточного увлажнения. Избыточная влажность, вызываемая весенним или осенним затоплением садов, неправильным орошением или же на участках сада с неровным рельефом местности и с близким залеганием грунтовых вод, угнетает ростовые процессы и резко снижает зимостойкость плодовых деревьев. При избыточной влажности почвы вследствие ухудшения аэрации и повреждения корневой системы подавляются ростовые процессы, наблюдается ограниченное поступление воды и минеральных веществ в растение. Поглотительная способность корней снижается в 2—3 раза по сравнению с корневыми системами деревьев, растущих в условиях оптимального увлажнения. При повреждении корневой системы в одних случаях отмирают только всасывающие корни или же понижается их поглотительная способность, в других — отмирают всасывающие корни и повреждаются паренхимные клетки проводящих тканей. В клетках последних часто образуется вещество типа индиго, имеющее сине-фиолетовую окраску, в третьих — происходит гибель корней и всего растения. Чем дольше плодовые растения находились в условиях избыточного увлажнения и чем выше температура окружающего воздуха, тем сильнее повреждается корневая система, ухудшается общее состояние деревьев и они погибают. Деревья, произрастающие на переувлажненных участках, позже трогаются в рост, медленно растут, листья на них мелкие, преждевременно опадают. При избыточной влажности почвы снижается восстановительная активность тканей, происходит смещение рН клеточного сока в кислую сторону, нарушается структурная организация протоплазмы. Вследствие переувлажнения почвы происходит увеличение количества и размеров 7. Ультраструктура хлоропластов листьев яблони сорта Мекинтош при оптимальном (а) и избыточном (б) увлажнении: Хп — хлоропласт, Гр — граны, Ко — клеточная оболочка, JI — фотосинтетические ламеллы. Пг — пластоглобулы, Т — тоно-пласт, Мх — митохондрия, Я — ядро (фото А. М. Силаевой, увеличение в 25 000 раз). 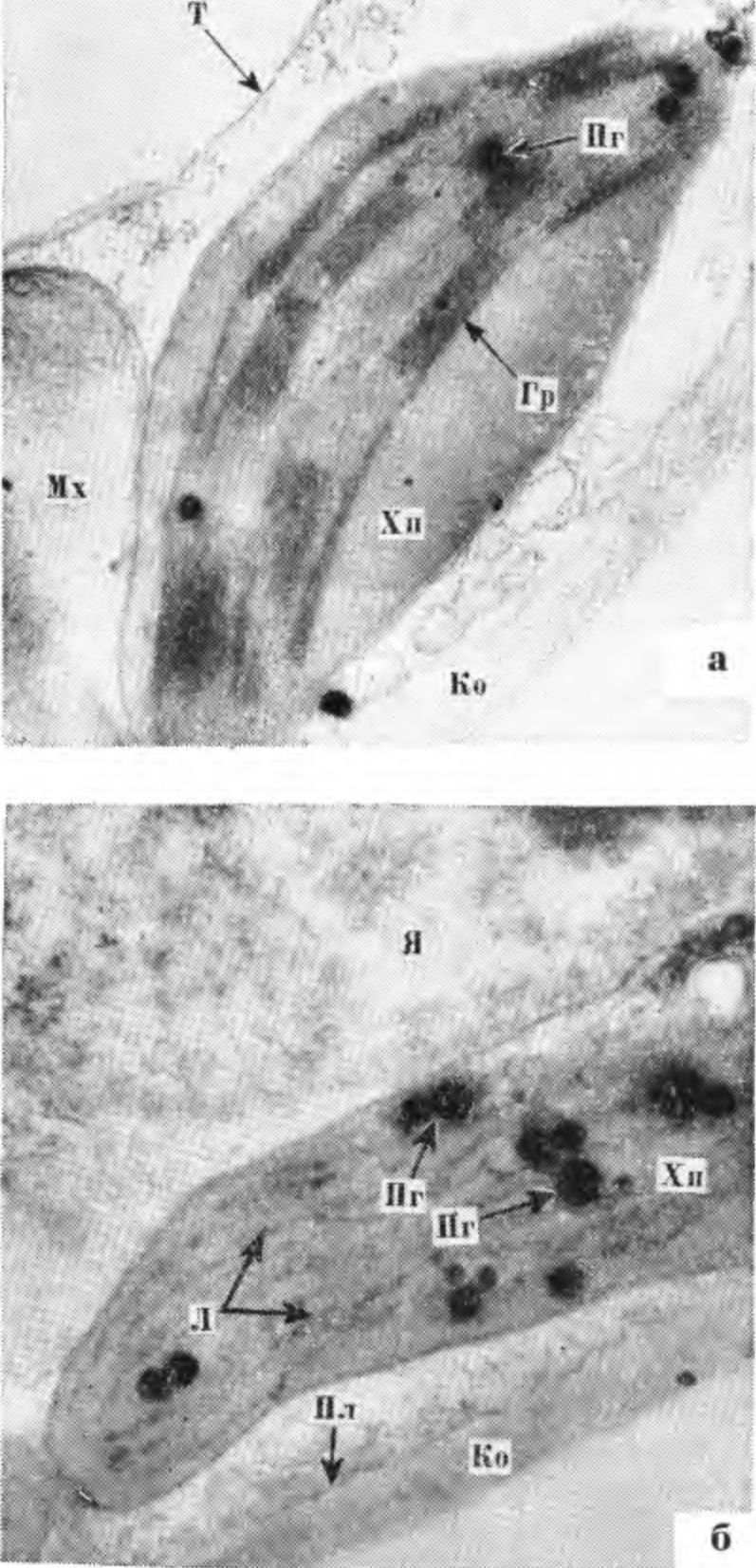
пластоглобул, значительно уменьшается количество фотосинтетических мембран и наблюдается деградация гранальной структуры хлоропластов (рис. 7). Изменяется состояние воды в клетке и снижается степень гидратации коллоидов, происходит резкое уменьшение запасов активной воды и интенсивное ее вымораживание в конце вегетационного периода в диапазоне температур от 0 до минус 5 °С (по данным ядерно-магнитного резонанса), возрастает водный дефицит, повышается осмотическое давление клеточного сока, происходит изменение состояния пла-стидного комплекса, уменьшается количество зеленых пигментов и появляется хлороз. При избыточной влажности почвы нарушается стабильность пластидного аппарата, разрушается хлорофилл прежде всего в молодых листьях верхней части побегов. При затоплении деревьев изменяются размеры орга-нелл — увеличиваются размеры митохондрий и уменьшаются — хлоропластов при деградации фотосинтетических мембран в последних (рис. 8). При повышенной влажности почвы инги-бируется также образование веществ фе-нольной природы, таких же как антоцианы и халконы, выполняющих защитную функцию в устойчивости растений к низким, переменным и высоким температурам. При избыточной влажности почвы резко подавляются ростовые процессы, ухудшается качество плодов, снижается урожайность и степень зимостойкости коры и камбия ствола, ветвей и развилок. Нередко такие деревья в последующие годы вымерзают полностью. Избыточное увлажнение вызывает глубокие и разносторонние нарушения физиологических функций плодовых растений. В связи с этим при закладке новых садов особое внимание необходимо уделять выбору и подготовке участка под сад, подбору устойчивых к избыточному увлажнению сортов и применению фосфорно-калийных удобрений, повышающих общую устойчивость деревьев и снижающих отрицательное влияние избыточного увлажнения. При орошении садов следует предусмотреть исключение возможного переувлажнения отдельных их участков. Наиболее устойчивым к избыточному увлажнению из районированных по Украине сортов яблони является Мекинтош. При ограниченном и переменном увлажнении в меньшей степени, чем при избыточной влажности, угнетаются ростовые процессы, подавляется образование поглощающих и развитие проводящих корней. Засухи, как и избыточное и неравномерное увлажнение, снижают степень зимостойкости коры, развилок ствола и ветвей плодовых деревьев. На коре образуются трещины или вдавленные пятна, иногда она вздувается и отстает. У деревьев, произрастающих в условиях ограниченной влажности почвы, всегда уменьшается листовая поверхность; листья изменяют окраску, хлорофилл в них разрушается прежде всего возле жилок. Вследствие разрушения хлорофилла ослабляется фотосинтез и наблюдается общее расстройство метаболизма из-за недостаточного образования органических веществ и дефицита энергетических резервов. Все листья с измененной окраской осыпаются среди лета. Наличие листьев с признаками водной недостаточности свидетельствует об опасном нарушении водного режима и необходимости полива. При недостатке азота в почве также изменяется окраска листьев равномерно по всей площади листа. Около жилок хлорофилл разрушается позже, чем по краям листа и между жилками. При продолжительном нарушении водного режима дерева изменяется не только окраска листьев, но и окраска побегов, ветвей и ствола, кора становится коричневато-желтой. Наличие побегов, ветвей и ствола с подобной окраской свидетельствует о том, что растения находятся в состоянии водного дефицита. Под воздействием засухи ускоряется процесс старения растений. Клетки и их коллоидные частицы дегидратируются, т. е. теряют способность удерживать воду. Дегидратация сопровождается понижением степени дисперсности коллоидов и изменением количества запасных веществ. При ограниченном и переменном увлажнении снижается интенсивность фотосинтеза и подавляется синтез сложных углеводов, нарушается белковый обмен и как результат нарушения функциональной деятельности организма снижается зимостойкость деревьев, прежде всего коры и камбия ствола, развилок и ветвей. Довольно эффективным способом предупреждения повреждения низкими и переменными температурами коры ствола и развилок у деревьев слабозимостойких сортов является двойная прививка, т. е. выращивание растений с так называемым промежуточным морозоустойчивым скелето-образователем. Такую прививку еще в тридцатые годы рекомендовали И. В. Мичурин и П. Г. Шитт. Широкие опыты по применению двойной прививки яблони и груши были проведены в Ленинградской области Н. Г. Жучковым, а в условиях Украины — И. К. Крывдой. И. А. Шереметом, А. Г. Ше-пельской, М. А. Соловьевой и др. Для яблони лучшим штамбообразовате-лем оказалась Боровинка, Антоновка обыкновенная, для груши — Александровка и некоторые формы груши лесной. Исследования, проведенные в Украинском НИИ садоводства и на Млеевской опытной станции садоводства им. Л. П. Симиренко, показали, что яблоня незимостойких сортов Ренет Симиренко, Джонатан, Кальвиль снеж- 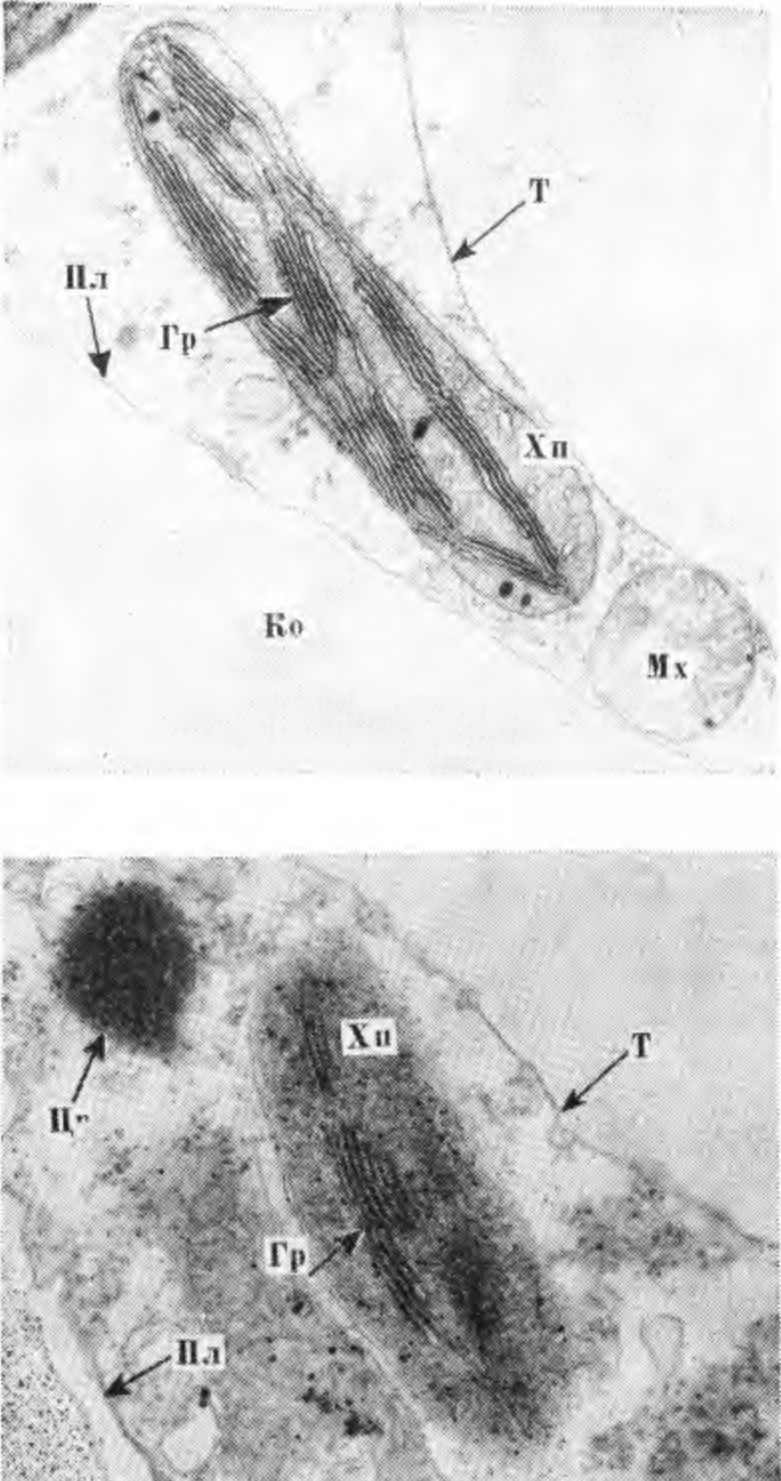 
8. Фрагменты клеток паренхимы коры побегов яблони сорта Мекинтош при оптимальной (вверху) и избыточной (внизу) влажности почвы: Хп — хлоропласт, Мх — митохондрия, Ко — клеточная оболочка, Цг — цитоглобула, Т — тонопласт, Пл — плазмалемма (фото А. М. Силаевой), (увеличение в 25 000 раз). ный и других, привитых в крону деревьев морозоустойчивых сортов Боровинка, Антоновка обыкновенная и других, сохранялись после суровых зим неповрежденными или повреждались в слабой степени, а привитые в корневую шейку — вымерзали полностью или повреждались в сильной степени. В отдельные зимы погибало 30—60 % и более деревьев, привитых в корневую шейку. Урожайность деревьев яблони, привитых в крону морозоустойчивых сортов, в связи с лучшей их сохранностью после суровых зим обычно на 10—20 или даже 25—35 % превышала урожайность привитых в корневую шейку. ПОВРЕЖДЕНИЕ КОРЫ У КОРНЕВОЙ ШЕИКИ Повреждение коры у корневой шейки наблюдается в садах у молодых деревьев яблони, груши, черешни, сливы и абрикоса в результате их окучивания, особенно в теплую дождливую осень, и у саженцев в прикопке. Такой же тип повреждения наблюдается в тех случаях, когда снег выпадает на неза-мерзшую влажную почву, корневая система не прекратила рост и в надземную часть интенсивно поступает вода. У корневой шейки или немного выше на коре могут появиться продольные трещины — одна, две и больше. Иногда трещины бывают неглубокие с одной стороны растения. Они быстро заживают. В большинстве же случаев трещины образуются глубокие в разных частях ствола и приводят к гибели растения. Гибель коры в этом случае является результатом неполного вызревания тканей ствола и недостаточного развития покровной ткани — перидермы, которая состоит из пробки, или феллемы, пробкового камбия, или феллогена, и фелло-дермы и является защитной тканью, предупреждающей повреждения морозами и солнечные перегревы тканевых элементов коры в разные периоды зимы (Степанов С. Н., 1981). В плодовых питомниках Украины, а также в молодых садах довольно часто наблюдается кольцевое повреждение ствола выше корневой шейки и повреждение тканей возле почек на стволе у однолетних саженцев в результате зимнего вымокания. В годы, когда зимой морозные дни чередуются с продолжительными оттепелями, в пониженных местах скапливается талая вода. Длительный застой ее и приводит к повреждению коры и камбия в различных участках ствола выше корневой шейки. Деревья с таким типом повреждения восстанавливаются только в тех случаях, когда кольцевое повреждение бывает выше места прививки и растения срезают на повторный рост. Отмирание коры и камбия на стволе саженцев, длительное время находившихся в воде, происходит вследствие изменения типа дыхания растений. В среде с ограниченным количеством кислорода дыхание становится анаэробным, происходит повышенный расход органического вещества, ткани растений быстро истощаются, одновременно образуются недоокисленные продукты, токсически действующие на растения. В отдельные годы наблюдается повреждение саженцев в питомнике ледяной коркой. В тех случаях, когда ледяная корка бывает продолжительный период и слой ее довольно толстый, она механически повреждает кору ствола саженцев у корневой шейки или на разном расстоянии от нее. Гибель тканей под ледяной коркой может произойти в результате увеличения в несколько раз содержания углекислого газа, который и отравляет ткани растений (Ракитина 3. Г., 1967). Различий в повреждении ледяной коркой стволов саженцев разных сортов не обнаружено. В зимнее время при резких колебаниях температуры и высокой влажности воздуха происходит оледенение ветвей, слой льда разной толщины сплошь покрывает ветви и стволы дерева. Перегруженные льдом ветви плодоносящих, ранее поврежденных морозом деревьев, иногда обламываются. При непродолжительном периоде оледенения механически повреждаются саженцы в питомниках, особенно в прикопке, и молодые деревья в садах. Плодоносящим же деревьям особого вреда оно может и не причинить. МОРОЗОБОИНЫ Морозобоины возникают вследствие длительного воздействия низких температур на дерево. Возникают они чаще на северной стороне дерева, хотя не редко бывают и в разных частях ствола. Повреждаются прежде всего молодые сильнорастущие деревья. Морозобоины — это повреждение коры и древесины в виде глубоких продольных трещин, доходящих иногда до центра дерева. Кора вдоль трещин, как правило, отстает от древесины, увеличивая этим размер раны. Такие повреждения наблюдаются при сильных морозах, особенно при резких колебаниях температуры в дневное и ночное время. Растрескивание стволов в большинстве случаев происходит в ночное время и сопровождается сильным треском. Морозобоины можно обнаружить не только на стволах, но и на толстых ветвях. Повреждение морозобоинами связано с продолжительностью ростовых процессов и со степенью вызревания древесины. Деревья с затянувшимся осенним ростом повреждаются сильнее, чем вовремя закончившие рост. Имеет значение также оводненность тканей растений и состояние воды в растениях. При замерзании объем ткани уменьшается, образуются трещины по мере того, как вода переходит в межклеточное пространство, а воздух вытесняется при образовании льда. Радиальные трещины на стволах вызываются тангентальными сокращениями древесины без ее радиального сокращения при воздействии мороза. Клетки разрушаются, разрываются или расслаиваются, когда внутриклеточные пространства слишком малы, чтобы ассимилировать весь образовавшийся лед. Степень повреждения различных сортов и деревьев одного сорта разного возраста неодинакова. Деревья летних сортов яблони обычно повреждаются слабее, чем осенних и особенно зимних. Более молодые деревья повреждаются сильнее, чем деревья, вступившие в пору плодоношения; высокоштамбовые — сильнее, чем низкоштамбовые. На переувлажненных участках сильно повреждаются не только молодые, но и плодоносящие деревья яблони, груши, черешни, сливы, вишни и абрикоса. Степень морозоустойчивости стволов зависит от физиологического состояния растений. При благоприятных условиях выращивания растений морозобоины могут зарасти, оставляя рубцы, которые при отсутствии повторных повреждений со временем становятся малозаметными. При заживании широких и глубоких морозобоин в результате активной деятельности неповрежденного камбия или разрастания паренхимных клеток флоэмных лучей возле поврежденных участков образуется каллюс. Со временем клетки каллюса преобразуются в клетки камбия с последующей дифференциацией флоэмы и ксилемы. При повторных повреждениях стволов и последующих заживаниях ран наблюдается как бы слоистое строение древесины стволов плодовых растений. В периферической зоне каллюса образуется перидерма. Ауксины активизируют процессы регенерации у поврежденных морозом деревьев. Если поврежденные морозом ткани очистить до здоровых, обработать раствором гетероауксина совместно с ланолином, петролатумом, садовой замазкой, содержащей гетероауксин, гиббереллином или КАНУ (0,25 — 0,5 %), то процессы регенерации (В. В. Грохольский, 1981) проходят более интенсивно, чем у необработанных растений. Обработанные раны зарастают за 1—2 года. Надземная часть молодых и взрослых деревьев может повреждаться морозами в результате опрыскивания садов под зиму минеральными эмульсиями, обмазывания рыбьим жиром и другими маслами против грызунов. Жир, проникая в ткани коры, вызывает нарушение процессов обмена веществ в растении и прежде всего процессов дыхания, в результате чего повреждаются живые клетки коры и камбиальной зоны ствола и ветвей. У последних, особенно возле почек, отмирают отдельные участки коры и камбия. Почки у таких растений не распускаются, а распустившиеся вскоре засыхают. В связи с этим в питомниках и молодых садах применять рыбий жир и минеральные растительные масла в борьбе с грызунами не следует. Опрыскивание минеральномасляны-ми эмульсиями под зиму также часто приводит к вымерзанию или сильному повреждению взрослых плодоносящих деревьев яблони. Известны случаи гибели взрослых деревьев яблони в зимнее время после обработки их с осени соляровым маслом или обмазки стволов подсолнечным маслом при полной со^ хранности необработанных деревьев на рядом расположенных участках. Нигрол или нигроловая замазка приводят к сильному повреждению обработанных участков коры. СОЛНЕЧНЫЕ ОЖОГИ Солнечные ожоги на коре ствола и ветвей плодовых деревьев наблюдаются не только в зимний и зимне-весенний, но и в летний периоды. Наиболее сильно поражаются стволы и ветви деревьев, произрастающих в условиях пониженной относительной влажности воздуха и ограниченной влажности почвы. Появление солнечных ожогов зимой, особенно во второй ее половине, является следствием чередования солнечных дней с морозными ночами и связано с локализованным выходом клеток коры из периода покоя, усилением жизнедеятельности тканей в дневное время и последующим повреждением как поверхностных, так и более глубоко лежащих тканевых элементов коры. Происходит необратимая агглютинация хлоропластов в некротированных клетках коры. С повышением температуры степень повреждения стволов и ветвей увеличивается. Солнечные ожоги появляются на южной и юго-западной сторонах ствола и ветвей в виде темных пятен различной формы и размера. Вначале они сохраняются в виде небольших локальных участков, которые впоследствии увеличиваются в размере, кора шелушится или растрескивается на отдельных поврежденных участках и отмирает. В зависимости от степени повреждения отмирают поверхностные ткани коры или вся кора до камбия. Со временем отмершая кора отстает, оголяя древесину. На поврежденных участках коры появляется возбудитель черного рака — гриб Schpaeropsis malorum. Гибель тканей от солнечных ожогов возможна и без образования льда в них, ткани могут быть убиты продуктами обмена при температуре, вызывающей распад хлорофилла. Избыточный солнечный свет, низкие, высокие и переменные температуры приводят к деструкции и полной гибели клеток в результате фотодинамического эффекта. Появление солнечных ожогов в конце весны — начале лета наблюдается при внесении высоких норм азотных удобрений, ухудшении водного режима растений или при повреждении морозом камбия и клеток камбиальной зоны. Солнечные ожоги коры ветвей и ствола возможны и при других функциональных нарушениях растений. Обычно повреждаются участки коры ствола и ветвей, хорошо освещенных солнцем. Светоустойчивость растительных тканей — величина непостоянная. Она изменяется в зависимости от условий выращивания. Существуют физиологическая и онто генетическая адаптация растений к солнечному свету. Изменение ралоложения хлоропластов в клетках коры в зимнее время, их отхождение от клеточных оболочек, скучивание, образование антоцианов являются защитным приспособлением растительного организма от избыточного солнечного света. Различные сорта той или иной породы не в одинаковой степени подвержены солнечным ожогам. Наиболее сильно повреждаются сорта яблони зимнего срока созревания, особенно Ренет Симиренко, Кальвиль снежный, Бойкен, Бельфлер желтый, Штет-тинское красное, Джонатан, Пепин лондонский, из осенних — Ренет ландсбергский, Пармен зимний золотой, Ренет Баумана, из летних — Папировка и другие, особенно в районах недостаточного увлажнения. Ожоги на деревьях груши наблюдаются значительно реже, чем на яблонях. Молодые деревья в год посадки больше повреждаются весенними ожогами, чем те, которые росли в саду в течение нескольких лет. Еще сильнее повреждаются пересаженные деревья, особенно те, что перед посадкой росли на неплодородных землях. Значительно сильнее подвержены солнечным ожогам сорта, сформированные в условиях влажного климата и произрастающие при ограниченной влажности почвы. Растения, выращиваемые в условиях недостаточного или неравномерного увлажнения, сильнее повреждаются, чем выращенные при постоянном равномерном увлажнении. Деревья, произрастающие на легких сухих почвах, бедных питательными веществами, повреждаются сильнее, чем на суглинистых и на почвах, богатых органическими веществами. У таких деревьев отмирает кора в средней части ствола, повреждается камбий и клетки камбиальной зоны, что приводит к угнетению ростовых процессов, ухудшению общего состояния растений и преждевременной их гибели. В результате солнечного перегрева у растений, выращиваемых в условиях ограниченного или переменного увлажнения при низкой относительной влажности воздуха, повреждаются и отмирают отдельные паренхимные клетки коры, хлоропласты, расположенные ближе к периферии, и другие тканевые элементы. Повреждение проводящей системы всегда приводит к появлению солнечных ожогов. Иногда наблюдается кольцевое повреждение всей коры, камбия и древесины не только в зимний и зимне-весенний периоды, но и в конце весны — начале лета. При кольцевых ожогах листья начинают увядать вскоре после их образования, затем они засыхают и к концу июня растение погибает. Ширина кольцевых ожогов бывает разной — от 1—2 до 7,5—10 см и более. На расстоянии 5—6 см от мертвой полосы кольцевого ожога, иногда выше или ниже появляются небольшие локализованные участки поврежденной коры, камбия и древесины, а в средней части ствола и выше наблюдается повреждение клеток вторичной флоэмы. В конце кольцевого повреждения гибнут все клетки коры, древесины и камбия. Сильно поврежденные ожогами деревья погибают в первой половине лета. Деревья со средним и слабым повреждением продолжают расти, под поврежденным слоем у них образуется каллюс, который позже дифференцируется с последующим образованием клеток флоэмы и ксилемы. При слабом повреждении восстановление проводящих тканей может произойти без образования каллю-са. В зависимости от степени повреждения изменяется и интенсивность ростовых процессов. В годы с резким колебанием температуры на коре ствола выше линии снежного покрова по всей длине штамба до основания скелетных и полу скелетных ветвей с южной или юго-западной стороны дерева или только на штамбе скелетных ветвей в результате ожога появляются различные пятна. К концу лета или в последующие годы кора отстает от древесины, оголяя ее. Оголившиеся слои древесины мертвеют. Чаще всего таким образом повреждаются незимостойкие сорта яблони и сливы Венгерка ажанская. У деревьев с поврежденными ветвями затрудняется передвижение питательных веществ и воды, нарушается нормальный ход физиологических процессов, снижается урожайность и последующая зимостойкость. Деревья могут погибнуть в год повреждения. Кора стволов повреждается солнечными ожогами при прикопке саженцев на зиму в сухую почву без обильного полива. К весне на стволе и скелетных ветвях отмирают отдельные участки коры, появляются темные сухие пятна в средней части ствола. Массовые солнечные ожоги саженцев в прикопке могут быть объяснены недостаточным поступлением воды в растение, слабой оводнен-ностью тканей и солнечным перегревом коры, разрушающие поверхностные и глубже лежащие ткани первичной и вторичной флоэмы и перидермы. Солнечные ожоги могут появляться при сильном перетягивании проволокой, плотными пленками, шпагатом и другими материалами, вызывающими кольцевание, а также при применении тонких полиэтиленовых пленок при защите стволов от грызунов (пленки ухудшают термический режим растений). Солнечные ожоги возникают и на плодах. Обычно когда после прохладной дождливой погоды наступают жаркие солнечные дни, на поверхности плодов появляются округлые бурые пятна, которые со временем темнеют, а затем загнивают. Плоды, пораженные солнечными ожогами, непригодны для хранения или употребления в свежем виде. Уменьшить вредное влияние солнечных ожогов можно путем подбора устойчивых к данному типу повреждения сортов плодовых деревьев, выращиванием растений всех основных плодовых пород с устойчивым скелетообразователем. В существующих посадках необходимо использовать различные приемы, уменьшающие солнечный перегрев растительных тканей зимой, весной и в начале лета. Хорошие результаты дает применение плотных полимерных пленок с отверстиями, винипластовых сеток, которые могут защищать стволы в течение первых 6—7 лет после посадки, обвязка различным обвязочным материалом (белой бумагой, стеблями подсолнечника, камышом и др.) и побелка водоэмульсионными красками типа ВС-511, ЭВА-27-А и др. (Грохольский В. В., 1981). ПОВРЕЖДЕНИЕ ПЛОДОВЫХ ПОЧЕК В годы с суровыми зимами или же с резкими колебаниями температуры во второй половине зимы наиболее сильно повреждаются плодовые почки у косточковых культур, несколько меньше у осенне-зимних сортов груши — реже повреждаются плодовые почки у яблони. Из косточковых культур к наименее устойчивым следует отнести цветковые почки у деревьев абрикоса, к более устойчивым, но все же частично повреждаемым — у черешен. У деревьев персика цветковые почки обычно более устойчивы, чем молодые ветви, на которых они расположены, хотя ветви персика менее морозоустойчивы, чем у абрикоса, и в годы с неблагоприятным сочетанием метеорологических факторов деревья персика повреждаются сильнее, чем абрикоса, черешни и других косточковых культур. Из всех косточковых пород наиболее морозоустойчивы плодовые почки вишни. При поражении листьев и ветвей грибными или вирусными болезнями морозоустойчивость плодовых почек и в целом деревьев резко снижается, из-за чего порядок в устойчивости различных плодовых пород может быть иным. В зависимости от длительности морозного периода наблюдается разная степень повреждения почек. В сильной степени поврежденные плодовые почки весной не раскрываются. Плодовые почки разных сортов одной породы повреждаются неодинаково. Наиболее заметные различия в повреждаемости наблюдаются у вишни, черешни, несколько меньше — у абрикоса и сливы. Почки у сливы сорта Анна Шпет, Венгерка обыкновенная и Ренклод Альтана повреждаются сильнее, чем у сортов Ренклод реформа, Венгерка итальянская и новых сортов селекции УНИИС и его опытных станций. Сильное повреждение плодовых почек наблюдается у вишен сортов Крупная чернявка, Подбельский, Анадольская; слабое — у Шпанки кра-снокутской, Гриота остгеймского, Самсонов-ки, Тургеневки, Любской. Из сортов абрикоса сильно повреждаются цветковые почки у сорта Краснощекий, у которого вымерзает от 90 до 100 % почек в зимы с резким колебанием температуры. У Колхозного, Аденис и ряда других новых селекционных сортов они более устойчивы к низким отрицательным температурам, морозоустойчивость цветковых почек у этих сортов выше на 2—4 °С, а при благоприятных условиях подготовки растений к зиме — на 4—6 °С выше, чем у основного районированного сорта Краснощекий. Наиболее часто плодовые почки повреждаются в конце зимы — начале весны. После завершения растениями глубокого покоя даже кратковременное потепление может вызвать начало их активной жизнедеятельности. Растения, вышедшие из периода покоя, повреждаются даже при незначительном похолодании. Плодовые почки абрикоса и черешни в южных районах Украины в зимнее время повреждаются чаще, чем в более северных, так как в южных районах продолжительные оттепели часто чередуются с внезапным похолоданием, что и приводит к гибели плодовых почек при сравнительно небольших морозах. Ростовые почки у большинства плодовых пород более устойчивы к низким температурам и реже вымерзают, чем плодовые, но у персика они часто бывают более устойчивы, чем ростовые. Разную степень устойчивости ростовых и плодовых почек можно объяснить разной водоудерживающей способностью и степенью дифференциации цветковых почек. Период покоя в плодовых почках заканчивается раньше, чем у ростовых. У них раньше начинаются и ростовые процессы. Известно, что с началом этих процессов морозоустойчивость отдельных органов и тканей резко снижается. Поврежденные плодовые почки буреют и, не раскрываясь, засыхают и опадают. При слабом повреждении весной они раскрываются медленно, иногда не полностью, а затем засыхают. В более поздние фазы их развития морозы повреждают чаще всего пестики — самую неморозоустойчивую часть цветка. Гибель пестиков является причиной снижения урожая в садах. На поперечных и продольных срезах поврежденные части почек или цветков имеют коричневую окраску. Степень морозоустойчивости плодовых почек подвержена значительным сезонным изменениям, которые, как показали исследования С. И. Елманова, Е. А. Яблонского, А. М. Шолохова (1966), зависят от прохождения последовательных этапов морфогенеза в годичном цикле развития. Наиболее высокой морозоустойчивостью цветковые почки у абрикоса и персика характеризуются в фазе мужского археспория. Чем продолжительнее этот период и чем позже он заканчивается, тем выше устойчивость почек к морозам. На последних этапах морфогенеза она прогрессивно снижается, достигая минимума в конце фазы двухъядерной пыльцы. Неодинаковой морозоустойчивостью характеризуются и разные зачатки цветков в одной почке. Устойчивость зачатков цветка, как и в целом цветковой почки, зависит от времени закладки и степени их дифференциации. Чем раньше заложились цветковые почки и чем сильнее они дифференцированы, тем больше опасность их вымерзания. Цветковые почки на более поздней стадии развития имеют большой процент клеток с мелкими вакуолями, а клетки с крупными вакуолями менее устойчивы, чем заполненные протоплазмой. Большое влияние на продолжительность и глубину периода покоя оказывает влажность почвы. Ограниченное и неравномерное увлажнение в течение вегетационного периода приводит к неполноценному осенне-зимнему покою, резко укорачивает его. В районах ограниченного увлажнения орошение является важным приемом повышения морозоустойчивости цветковых почек у косточковых культур. Положительные температуры плюс 15— 18 °С в зимнее время ускоряют развитие цветковых почек. В дальнейшем растения приобретают циклический тип развития, при котором на дереве одновременно имеются цветки, молодая завязь и созревающие плоды. Такие деревья имеют слабый прирост побегов. Облиственность их в 4—4,2 раза меньше, чем у обычных деревьев, урожайность резко снижается. Критическая температура гибели цветковых почек как разных сортов, так одного и того же сорта не постоянна. При резких колебаниях температуры во второй половине зимы или при наступлении ранних морозов в осенне-зимний период повреждаемость цветковых почек повышается. Существенные различия в морозоустойчивости цветковых почек отмечены по длине однолетних ветвей, а также в разных ярусах кроны дерева. В нижней части кроны почки повреждаются в сильной степени или полностью погибают, а в верхней значительная часть их сохраняется. В качестве примера можно привести данные повреждаемости цветковых почек в зиму 1966/67 г. у черешни сортов Китаевская черная, Любимица Дуки. У первого сорта в нижней части кроны сохранились здоровыми всего 0,3 % цветковых почек, а в верхней — 50,3, у второго — соответственно 5,8 и 36,8 %; урожай у сорта Китаевская черная в нижней части кроны полностью отсутствовал, а в верхней было собрано по 30 кг нормально развитых плодов с каждого дерева, у второго сорта урожай в нижней части дерева также отсутствовал, а в верхней составлял 11,1 кг с дерева. Аналогичные данные получены в зиму 1986/87 г. В зиму 1978/79 г. наблюдалась аналогичная зависимость. В нижней части кроны на расстоянии 2—2,5 м от поверхности почвы цветковые почки у сорта Любимица Дуки погибли полностью, а в верхней сохранились здоровыми 20 % почек, у Красавицы Киева сохранилось живыми соответственно 18,5 и 48; у Китаевской черной — 23 и 58 %. Подобная зависимость отмечена также у сливы сортов Венгерка обыкновенная и Венгерка итальянская. У Ренклода Альтана различий в повреждаемости морозами цветковых почек в разных ярусах кроны не было отмечено. У вишни Подбельская в верхней части на одних участках сохранилось 62,3 %, на других — 16 % цветковых почек, а в нижней — соответственно 3,6 и 2 %. Аналогичная зависимость наблюдалась у Шпанки ранней и других сортов. Наличие описанных различий можно объяснить состоянием апикальной меристемы, локализацией отдельных фенольных соединений и ассимилятов, оводненностью тканей в разных частях дерева, а также изменением пигментной системы в связи с интенсивностью освещения. Известно, что интенсивность света, прошедшего через толщину одного листа у многолетних древесных растений, составляет от 1/4 до 1/10 исходной интенсивности прямого света и соответственно оказывает влияние не только на величину, но и на направленность фотосинтеза, формирование фотосинтегического аппарата, изменение ультраструктурной организации органелл и состояние в них пигментов (Оканенко А. С., 1969). По ярусам кроны изменяется интенсивность и продуктивность фотосинтеза, реализуемая в синтезе ассимилятов, значительная часть которых является защитными веществами, обусловливающими степень подготовки растений к зиме и зимостойкость как цветковых почек, так и всего растения. Роль освещенности листового аппарата для формирования зимостойкости цветковых почек у косточковых, как и у других многолетних древесных плодовых растений, велика. М. Д. Кушниренко (1967) отмечает различия в оводненности тканей, в морфологических и структурных изменениях отдельных органов в разных ярусах дерева, предопределяющие не только степень морозоустойчивости генеративных и вегетативных органов плодовых растений, но и их засухоустойчивость. Следует отметить, что указанная выше зависимость в повреждаемости цветковых почек по разным ярусам кроны наблюдается не всегда. После засушливого лета в суровые зимы при сильных январских и февральских морозах в верхней части кроны обычно наблюдается более сильная повреждаемость и гибель цветковых почек и ветвей разного возраста, чем в средней и нижней частях. Особенно резко снижается зимостойкость цветковых почек у косточковых культур после дождливой и относительно теплой осени. В качестве примера можно привести данные гибели цветковых почек у косточковых в зиму 1978/79 г. при температуре минус 26— 30 °С и минус 28—32 °С. В районах с такими значениями минимальной температуры она составила у вишни 42 и 82 %, сливы — 73 и 97, черешни — 54 и 100, абрикоса — 100 %. Диапазон колебания критических температур в разные годы для одного и того же сорта находится в пределах 4—6 и даже 10—14 °С. Резко снижается зимостойкость цветковых почек в годы, когда суровым зимам предшествуют летние засухи или же растения выращивали в условиях избыточного увлажнения, ограниченного или повышенного уровня азотного питания. Повреждаемость цветковых почек в значительной степени зависит от адаптационных свойств. Процессы восстановления поврежденных морозом растений определяются их регенерационной способностью. При регенерации, как известно, изменяется направленность процессов метаболизма, усиливается синтез ДНК, белков, происходит локализация ауксинов. Последние усиливают регенерацию. В отдельных случаях она осуществляется путем преобразования паренхимных тканей в эмбриональные. При повреждении морозами генеративных почек вишни А. И. Евстратов (1964, 1975) наблюдал тип регенерации, при котором происходит пробуждение спящих почек в пазухах кроющих чешуй и осуществляется регенерация дополнительных конусов нарастания. В местах их повреждения возникает каллусный наплыв. Из вторичных меристематических тканей формируется новая зачаточная почка. Этот тип регенерации может быть использован в исследованиях с тканевыми культурами. В целях уменьшения опасности вымерзания плодовых почек абрикоса в сороковых годах П. Г. Шитт предложил летнюю обрезку, способствующую усилению вегетативного роста и перемещению плодоношения с укороченных плодовых веточек на побеги вторичного летнего прироста. При помощи этого приема сдвигаются сроки закладки плодовых почек, окончания периода покоя и распускания почек. Одновременно уменьшается возможность повреждения плодовых почек в зимний и ранневесенний периоды. В годы с сильными морозами обрезка черешни, как и других плодовых деревьев, должна проводиться в соответствии со степенью повреждения цветковых почек в разных ярусах кроны дерева. В настоящее время доказано положительное влияние летней обрезки на уменьшение повреждения плодовых почек морозами и на изменение сроков цветения деревьев абрикоса. Так, по данным С. Елманова и А. Щербины, в зиму 1947/48 г. на обрезанных по методу П. Г. Шитта деревьях вымерзло 32 % почек, а на контрольных (необрезанных) — 78 %. Обрезанные деревья зацвели на 4—7 дней позднее, чем контрольные. Уменьшение повреждения плодовых почек на обрезанных деревьях наблюдалось также в опытах И. М. Рядновой, К. Г. Никишина (1949) и др. По данным У. X. Чендлера (1960), обрезка задерживает развитие плодовых почек у персика и уменьшает их гибель. В. Г. Гарднер, Ф. Ч. Бредфорд, Г. Д. Гукер (1934) сообщают, что в районе Брэндевнль (штат Мус-сури) гибель плодовых почек на обрезанных деревьях персика составила 81,9 %, на необрезанных 98 %. Сохранившиеся почки на обрезанных деревьях обеспечили полный урожай, а на необрезанных были только единичные плоды. Сроки закладки цветковых почек у яблони, по данным Н. Н. Пилипенко (1959), сильно варьируют и зависят от комплекса агротехнических приемов. На срок их закладки большое влияние оказывают водный и питательный режимы растения. Под влиянием засухи плодовые почки закладываются раньше, под влиянием обрезки и удобрений — позже. Так, в опытах Н. Н. Пилипенко у необрезанных деревьев яблони сорта Папировка начало закладки плодовых почек было отмечено 3 июля, на сильно обрезанных — 11 июля, а при подсушивании почвы — 28 июня. Повреждение плодовых почек зависит от общего состояния дерева, интенсивности роста и облиственности. Плодовые почки, сформировавшиеся на деревьях, очень рано закончивших рост, отличаются низкой морозоустойчивостью. На мощных, хорошо развитых ветвях плодовые почки более устойчивы, чем на тонких, коротких, которые рано заканчивают рост и бывают слабооблиственными или же имеют поврежденные листья. Наиболее морозоустойчивые плодовые почки у основания однолетних побегов. Проведенный анализ плодовых почек вишни, сливы и черешни показал, что повреждение 40—50 и даже 60—65 % плодовых почек при хорошей их закладке не приводит к снижению урожая — оставшееся количество здоровых почек обеспечивает нормальный урожай. Применение азотных удобрений и сплошное опрыскивание деревьев известью снижают гибель почек абрикоса (Никишин Г. К., 1949; Васильев И. М., 1956). В опытах Г. К. Никишина после мороза в 16,1 °С неопрысну-тые известью деревья потеряли 65 % плодовых почек, а опрыснутые — всего 25 %. Урожай был соответственно 70—80 и 110—120 кг с дерева. Как показали исследования отдела физиологии и зимостойкости УНИИС, криопро-текторы, в частности полиэтиленоксиды (Г1ЭО) молекулярной массы 400 и 1500, способствуют повышению морозоустойчивости цветковых почек у деревьев абрикоса при обработке их в осенний период (Соловьева М. А., 1984). Сохранность цветковых почек при обработке растений водными растворами ПЭО-1500 в концентрации 1 —1,5 %, а также смесью ПЭО-400 и ПЭО-1500 была выше на 63—65 %, чем у необработанных растений, а у сорта Дарунок Малаховой цветковых почек сохранилось в 2 раза больше, чем у контрольных. В регуляции ростовых процессов особая роль принадлежит биологически активным веществам. В последнее время в садоводстве довольно широко используются синтетические биологически активные вещества, регулирующие рост, развитие и урожайность плодовых растений, так называемые экзогенные регуляторы роста. Среди них особое место занимают такие ретарданты, как ССС (хлорхолинхлорид — 2-хлорэтин — триме-тиламмоний хлористый, хлормекват) и его аналог тур, а также производные димети-ламиноянтарной кислоты (диминозид, алар, В-9, дяк) и др. Большинство из экзогенных регуляторов роста способствует ускорению прохождения отдельных этапов онтогенеза, обусловливающих ускорение перехода к генеративному развитию и более раннему вступлению в пору плодоношения плодовых деревьев. Они ингибируют линейный рост и стимулируют камбиальную деятельность (Кефели В. И., 1974), оказывают влияние на оводненность тканей, биосинтез пигментов, углеводов, некоторых фенольных соединений и степень устойчивости к низким и переменным температурам. В зависимости от применяемых концентраций экзогенных регуляторов роста, их химической природы и генотип и ческой специфичности сорта наблюдаются различия в эффективности их действия. Как показали исследования отдела физиологии и зимостойкости УНИИС (Соловьева М А., 1984), алар и его аналог дяк более универсального действия, чем ССС или тур. Первые ингибируют линейный рост и стимулируют камбиальную деятельность у саженцев в питомниках, молодых и взрослых плодоносящих деревьев яблони в садах, также стимулируют закладку цветковых почек. Урожайность плодоносящих деревьев в условиях сада увеличивается на 25—30 %, в контролируемых условиях она возрастает в 1,3— 2 и более раза. Под влиянием препарата дяк изменяется угол отхождения ветвей: он становится более тупым, сокращается расстояние междоузлий и наблюдается более раннее окончание ростовых процессов. При применении препарата дяк в концентрации 0,2 % в ранневесенний период через 10—20 дней после цветения у растений повышается степень морозоустойчивости камбия и клеток камбиальной зоны. При более высоких концентрациях препарата отмечена тенденция к снижению морозоустойчивости растений. ССС или тур сильнее ингибируют линейный рост у саженцев в питомниках и молодых деревьев яблони в контролируемых условиях; в саду на взрослых плодоносящих деревьях яблони сортов Кальвиль снежный, Мекинтош, Слава победителю и других не всегда подавляют линейный рост и способствуют повышению урожайности в районах Полесья УССР. Эти препараты применяются в концентрациях 0,5 и 0,6 %. У молодых деревьев в первый год линейный рост инги-бируется, а затем после прекращения обработки он заметно стимулируется. Количество плодов на второй год применения препаратов у молодых деревьев возрастало в 1,8— 4,3 раза по сравнению с необработанными. При ежегодном применении ССС или тура в концентрации 0,8—1 % или при двукратной обработке в концентрации 0,4 % плодоношение деревьев этих сортов становится периодичным. Препараты тур и ССС часто вызывают краевой хлороз листьев, сохраняющийся в отдельных сортов яблони до осени, а также наблюдается резкое сокращение междоузлий у обработанных деревьев. При повышенных концентрациях препарата тур (0,8—1 %) морозоустойчивость древесины, верхней части ветвей прошлого года снижается, особенно во второй половине зимы. Экзогенные регуляторы роста эффективны только на фоне высокой агротехники и при сохранении листового аппарата от повреждений вредителями и болезнями. Ингибирующее влияние на ростовые процессы и на гибель точек роста отдельных тканей и даже целых растений оказывают гербициды, которые широко используются в садах для подавления развития и уничтожения сорняков. Для борьбы со злаковыми сорняками в садах применяют гербицид строго избирательного действия — симазин в разных концентрациях, а также 2,2-дихлор-прогшонат Na (далапон), трихлорацитат Na (ТХА) и другие уничтожающие как однолетние, так и многолетние сорняки из семейства злаковых. Проведенными исследованиями в отделе физиологии и зимостойкости УНИИС по изучению влияния гербицидов на морозоустойчивость деревьев яблони в опытах А. И. Мартыненко показано, что в оптимальных нормах, рекомендованных отделом агротехники для применения в садах симазина 4 (кг/га), морозоустойчивость деревьев яблони не изменяется. С увеличением дозы симазина до 6—7,5 кг/га морозоустойчивость деревьев яблони заметно снижается. В районах с резко континентальным климатом способ использования гербицидов и их концентрации должны соответствовать степени морозоустойчивости плодовых деревьев и исключать возможность снижения последней. При неосторожном применении гербицидов, когда последние попадают на листья и плоды, поражают их, листья снижают фотосинтетическую активность, раньше опадают, плоды приобретают уродливую форму, морозоустойчивость таких плодовых деревьев резко снижается. |